 E-submission
E-submission
Articles
- Page Path
- HOME > J Pathol Transl Med > Volume 46(2); 2012 > Article
-
Original Article
Expression of HAT1 and HDAC1, 2, 3 in Diffuse Large B-Cell Lymphomas, Peripheral T-Cell Lymphomas, and NK/T-Cell Lymphomas - Soo Kee Min, Young Ho Koh1, Yunwoong Park1, Hyo Jung Kim2, Jinwon Seo, Hye-Rim Park, Seong Jin Cho3, In Sun Kim4
-
Korean Journal of Pathology 2012;46(2):142-150.
DOI: https://doi.org/10.4132/KoreanJPathol.2012.46.2.142
Published online: April 25, 2012
Department of Pathology, Hallym University Sacred Heart Hospital, Hallym University College of Medicine, Anyang, Korea.
1Ilsong Institute of Life Science, Hallym University, Anyang, Korea.
2Department of Hemato-Oncology, Hallym University Sacred Heart Hospital, Hallym University College of Medicine, Anyang, Korea.
3Department of Pathology, Hallym University Kangdong Sacred Heart Hospital, Hallym University College of Medicine, Seoul, Korea.
4Department of Pathology, Korea University College of Medicine, Seoul, Korea.
- Corresponding Author: Seong Jin Cho, M.D. Department of Pathology, Hallym University Kangdong Sacred Heart Hospital, 150 Seongan-ro, Gangdong-gu, Seoul 134-701, Korea. Tel: +82-2-2224-2557, Fax: +82-2-2224-2214, apilas@hanmail.net
© 2012 The Korean Society of Pathologists/The Korean Society for Cytopathology
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Abstract
-
Background
- It has generally been proven that histone acetylation and deacetylation are involved in the malignant transformation. To date, however, this has rarely been studied in cases of malignant lymphoma.
-
Methods
- We studied nine cases of reactive lymphoid hyperplasia, 78 cases of diffuse large B-cell lymphoma (DLBCL), 13 cases of peripheral T-cell lymphoma, not otherwise specified (PTCL-NOS), and 13 cases of extranodal NK/T-cell lymphoma, nasal type (NKTCL). Thus, we attempted to elucidate the associations of the degree of the expression of histone acetyltransferase 1 (HAT1), histone deacetylase (HDAC) 1, HDAC2, and HDAC3 with the clinical behaviors of above malignant lymphomas using the immunohistochemistry and a western blot analysis.
-
Results
- The degree of the expression of HAT1 was higher in cases of DLBCL, PTCL-NOS or NKTCL as compared with reactive lymphoid hyperplasia (p<0.05). The degree of the expression of HAT1 was correlated with that of HDAC1 in cases of DLBCL or NKTCL (p<0.05). The degree of the expression of HAT1 and HDAC1 was correlated with a poor survival in cases of DLBCL or PTCL-NOS (p>0.05).
-
Conclusions
- HAT1, HDAC1, and HDAC2 play a critical role in the development of malignant lymphomas. Both HAT1 and HDAC1 might be indicators for a poor prognosis in cases of DLBCL as cooperating factors.
- Materials
- This study was approved by the Institutional Ethics Committee of our medical institution. We selected nine cases of RLH, 78 cases of DLBCL, 13 cases of PTCL-NOS, and 13 cases of NKTCL based on the criteria that the paraffin blocks were well preserved with a sufficient amount of tissue for evaluation. The clinical data, pathology reports and pathology slides were reviewed. In addition, 3-mm tissue microarrays were made.
- Immunohistochemical (IHC) stains
- IHC reactions were performed on paraffin tissue sections using an automated IHC stainer (Ventana BenchMark XT, Ventana Medical Systems Inc., Tucson, AZ, USA) according to the manufacturer's protocol. Detection was done using the Ventana i VEIW DAB detection kit (Ventana Medical Systems Inc.). Briefly, IHC staining was performed as follows: 4-µm-tissue sections were deparaffinized using EZ Prep solution. CC1 standard (pH 8.4 buffer containing Tris/Borate/ethylenediaminetetraacetic acid) was used for antigen retrieval at 99℃ for 60 minutes. i VIEW inhibitor (3% H2O2, endogenous peroxidase) was blocked at 37℃ for four minutes. Slides were incubated with primary antibodies (Table 1) at 42℃ for 32 minutes and secondary antibody to i VIEW biotinylated Ig at 37℃ for 8 minutes. Slides were incubated in i VIEW streptavidin HRP at 37℃ for 8 minutes and then DAB+H2O2 substrate for 8 minutes, which was followed by counterstaining with hematoxylin and bluing reagent at 37℃. Reaction buffer (pH 7.6 Tris buffer) was used as a washing solution.
- Interpretation of IHCs
- Antibodies to HAT1, HDAC1, HDAC2, HDAC3, and Ki-67 stained the nuclei. The IHCs associated with histone-related proteins (HAT1, HDAC1, HDAC2, and HDAC3) were interpreted based on the intensity: 0, 1+, 2+, and 3+, where 0 is negative, but Ki-67 based on the proportion (< 26%, 26-50%, 51-75%, and>75%). In HAT1 and HDAC1 with an intensity of 1+, the intensity of nearby endothelial cells was served as the control one. In HAT1 and HDAC1 with an intensity of 3+, however, the intensity of germinal center cells was served as the control one. Besides, in HDAC2 and HDAC3 with an intensity of 1+, the intensity of endothelial cells was served as the control one. In HDAC2 and HDAC3 with an intensity of 3+, however, the intensity of nearby histiocytes was served as the control one. Furthermore, 2+ was considered the moderate intensity between 1+ and 3+ (Figs. 1, 2). Finally, the intensity and proportion of histone-related proteins were compared using a Kaplan-Meier survival and a Spearman correlation coefficient.
- Western blot
- Western blot analysis was available only for three cases of RLH, four cases of DLBCL and three cases of PTCL-NOS. Protein extraction was performed with the modified methods of Azimzadeh et al.11 Briefly, western blot analysis was performed as follows: paraffin sections were prepared from the samples of RLH and malignant lymphomas by the microdissection. This was followed by processing for western blot analysis. Then, the paraffin sections were subjected to deparaffinization after incubated twice with xylene at room temperature for ten minutes. Following the dehydration in a graded series of ethanol (100%, 90%, and 70% EtOH) for ten min each, the sections were washed with a 0.5% octylglucoside (Sigma-Aldrich, St. Louis, MO, USA). The deparaffinized sections were resuspended in an extraction buffer (20 mM Tris-HCl, pH 8.8, 2% sodium dodecyl sulfate [SDS], 1% octylglucoside, 200 mM dithiothreitol, 200 mM glycine; Sigma-Aldrich) and then sonicated three times at 24℃ for 30 seconds in Brasonic 8510R-DTH sonicator (Brason, Danbury, CT, USA). This was followed by a 10-minute boiling at 100℃ and a centrifugation at 15,000 ×g at 4℃ for 10 minutes. Then the protein in the supernatant was separated by the SDS-polyacrylamide gel electrophoresis and then transferred onto a Hybond-N nitrocellulose membrane (GE Healthcare, Pittsburgh, PA, USA). Monoclonal anti-actin antibodies were served as loading controls. In addition, rabbit polyclonal anti-HAT1 antibodies and monoclonal anti-HDAC1 ones were used to detect HAT1 and HDAC1 (Table 1).
- Interpretation of western blot
- The intensity of anti-actin band and anti-HAT1/HDAC1 one was semi-automatically measured using the "wander tools" and "histogram" functions in the Adobe Photoshop (Adobe, San Jose, CA, USA) as previously described by Park et al.12 Then, the intensity was compared using one-way analysis of variance (ANOVA) with the Duncan's test in the SAS program (SAS Institute, Cary, NC, USA).
MATERIALS AND METHODS
- The mean age of patients was 61 years old in 78 cases of DLBCL, 50 years old in 13 cases of PTCL-NOS and 58 years old in 13 cases of NKTCL. No male predilection or female one was observed in our series of clinical cases. The mean follow-up periods were 31, 18, and 15.8 months in cases of DLBCL, PTCL-NOS, and NKTCL, respectively (range, 1 to 162 months). Of the three types of lymphoma, the PTCL-NOS showed the poorest prognosis based on the death rate (Table 2).
- IHC studies
- The frequency of Ki-67 staining was the highest in the area of >26% and<50% and the lowest in that of >75% in all the three types of lymphoma. In cases of RLH, the intensity was mostly 2+ for HAT1, HDAC1 and HDAC2 but 0 and 1+ for HDAC3. Besides, in cases of DLBCL, PTCL-NOS, and NKTCL, it was mostly 3+ for HAT1, HDAC1, and HDAC2 but 0 and 1+ for HDAC3. The degree of the expression of HAT1 was significantly higher in cases of DLBCL, PTCL-NOS or NKTCL compared with RLH (p<0.05). The degree of the expression of HDAC1 was significantly higher in cases of DLBCL as compared with RLH (p<0.05). Besides, the degree of the expression of HDAC2 was significantly higher in cases of PTCL-NOS and NKTCL as compared with RLH (p<0.05) (Fig. 3). The proliferation index (Ki-67) was significantly correlated with the degree of the expression of HAT1 and HDAC1 in cases of DLBCL and that of HDAC2 in cases of NKTCL (p<0.05). In addition, it was also significantly correlated with the degree of the expression of HAT1 and that of HDAC1 in cases of DLBCL and NKTCL (p<0.05). Furthermore, there were significant correlations between the degree of the expression of HDAC1 and that of HDAC2 in cases of PTCL-NOS and between that of HDAC2 and that of HDAC3 in cases of DLBCL (p<0.05) (Table 3).
- Western blot analysis
- We lost many cases while preparing for a western blot analysis of the formalin-fixed, paraffin-embedded tissue samples. Besides, we also had no sufficient number of remaining cases that are available for the current study. Despite a small number of remaining cases, however, we have obtained the significant results as shown below: the density of protein band was reduced because the western blot analysis showed very strong protein bands for HAT1. This indicates that the density of protein bands was negative for cases of RLH. But the weak density was observed in case 4 as shown in Fig. 4A. The degree of the expression of HAT1 was significantly higher in cases of DLBCL (3/4) and those of PTCL-NOS (2/3) as compared with those of RLH (p<0.05). In the remaining cases of DLBCL (1/4) or PTCL-NOS (1/3), however, the degree of the expression of HAT1 was higher as compared with those of RLH. But this difference did not reach a statistical significance (Fig. 4). In addition, there were no any significant results in case of HDAC1.
- Survival rate
- Following an analysis of the survival curve, the degree of the expression of HAT1 and HDAC1 was higher in association with a poorer survival in cases of DLBCL and those of PTCL-NOS. But this did not reach a statistical significance (p>0.05). In addition, the degree of the expression of HDAC3 was also higher in association with a better survival in cases of NKTCL. But this also did not reach a statistical significance (p>0.05) (Fig. 5).
RESULTS
- It is well established not only that the carcinogenesis is commonly associated with the alteration of DNA sequences but also that epigenetic alterations are found in various types of cancers. Aberrant methylation of CpG islands in promoter regions causes the silencing of tumor suppressor genes in some cancers including malignant lymphoma.13,14 The imbalance between acetylation and deacetylation of histone protein induces the development, invasion and metastasis of the tumor. Increased histone acetylation may lead to the development of malignant lymphoma.2 Yasui et al.4 maintained, however, that the decrease of histone acetylation is involved not only in the development of tumor, but also its invasion and metastasis. The histone hypoacetylation is induced by a decrease in the activity of HAT or an increase in that of HDAC. The altered expression of HDAC proteins has also been reported to occur in cases of tumor. These alterations include the increased expression of HDAC1 in cases of gastric,15 prostate,16 colon,17,18 or breast cancer.19 Besides, it has also been disclosed that the increased expression of HDAC2 was present in cases of cervical,20 gastric,21 and colorectal cancer.20,22 A higher degree of the expression of HDAC3 was observed in cases of colon cancer.23 According to studies about hematopoietic malignancies, the degree of the expression of such histone-related proteins as HDAC1, HDAC2, and HDAC6 was significantly higher in cases of DLBCL or PTCL than normal lymphoid tissue.24 In addition, Marquard et al.25 maintained that a high degree of the expression of HDAC2 is more commonly seen in cases of aggressive CTCL rather than indolent cases. These authors also noted that the degree of the expression of HDAC6 is associated with a favorable outcome.25 Following the treatment with rituximab in cases of B-cell lymphoma, the degree of the expression of HDAC was associated with down-regulation of CD20 expression. The expression of CD20 mRNA and protein was repressed by recruitment of a histone deacetylase protein complex to the CD20 gene promoter.26 In addition, Agata et al.2 reported that histone acetylation determines an accessibility to the recombination of T-cell receptor γ-chain genes that play a direct role in executing a developmental switch in cell fate determination.
- Our results showed that the degree of the expression of HAT1 was significantly higher in cases of DLBCL, PTCL-NOS or NKTCL as compared with those of RLH (p<0.05). This is closely associated with the role of histone acetylation in inducing the expression of loose chromatin and thereby promoting the transcription. Epigenetics may play a secondary role in the development of tumor. It can therefore be inferred that the effects of histone acetylation would depend on the target oncogenes. The hypoacetylation of oncogenes would lead to the decreased oncogenesis. But opposite results are expected in tumor suppressor genes. Our results showed no specific oncogenes that play a critical role in the malignant transformation. In the current study, however, histone acetylation plays a critical role in the development of such cancers as DLBCL, PTCL-NOS, and NKTCL.
- On the other hand, many studies have examined the effects of HDAC inhibitors. In addition, the indications of SAHA have been approved for the treatment of CTCL by the US FDA. It has been known that HDAC inhibitors are involved in cell cycle arrest, differentiation and apoptosis in tumor cells. Some authors argue that isotype-specific HDAC inhibitors may be a more effective and safe agent that causes less adverse effects. Nevertheless, isotype-specific HDAC inhibitors are not popular up to present.27
- In the current study, the degree of the expression of HDAC1 was significantly higher in cases of DLBCL and those of HDAC2 in cases of PTCL-NOS and NKTCL as compared with RLH (p<0.05). This is in agreement with previous reports.24 Based on our results, it can be inferred that HDACs are involved in the expression of dense chromatin and this is associated with the repression of certain types of tumor suppressor genes, even though we did not disclose what these tumor suppressor genes are. HDAC1 plays a critical role in cases of DLBCL. But HDAC2 plays a critical role in cases of PTCL-NOS or NKTCL. This suggests that HDAC1- and HDAC2-specific inhibitors would be more effective for the treatment of DLBCL and that of PTCL-NOS and NKTCL, respectively.
- On the other hand, we could not verify the relationship between the expression of HDAC1 and HDAC2 and the survival of patients. Our results showed, however, not only that the degree of the expression of HAT1 and HDAC1 was significantly higher in relation to a poorer survival in cases of DLBCL or PTCL-NOS but also that the degree of the expression of HDAC3 was significantly lower in relation to a poorer survival in cases of NKTCL (p>0.05).
- In addition, the expressions of HAT1 were correlated with HDAC1 in cases of DLBCL (p<0.05). The proliferation index was significantly correlated with the degree of the expression of HAT1 and HDAC1 in cases of DLBCL (p<0.05). These results indicate that HAT1 and HDAC1 have a synergistic effect in development of DLBCL, both of which are involved in its aggressiveness as cooperating factors.
- Our results are also in agreement with the reports that both HATs and HDACs are involved in the development of cancer through a multi-step process together with several cooperating factors. In other words, HDACs act on specific genome regions by recruitment of DNA binding factors including transcription factors, nuclear receptors and epigenetic modifiers (methyl binding proteins, DNA methyltransferases, and histone methyltransferases). Considering these factors further specific studies are warranted to clarify the accurate mechanisms by which lymphoreticular malignancies occur. This will also be helpful for elucidating the effects of HDAC inhibitors against malignant lymphomas.
- In summary, our results are as follows:
-
The degree of the expression of HAT1 was higher in cases of DLBCL, PTCL-NOS or NKTCL as compared with RLH.
Presumably, HAT1 might act on some types of oncogene and thereby contribute to the development of such cancers as DLBCL, PTCL-NOS, and NKTCL.
The degree of the expression of HDAC1 and HDAC2 was higher in cases of DLBCL, PTCL-NOS, and NKTCL as compared with RLH. Both HDAC1 and HDAC2 are involved in the repression of certain types of tumor suppressor genes.
- In conclusion, both histone acetylation and deacetylation play a critical role in the development of such cancers as DLBCL, PTCL-NOS, and NKTCL. In addition, it can also be concluded that both HAT1 and HDAC1 might be indicators for a poor prognosis in cases of DLBCL, which is based on the following results:
-
HAT1 and HDAC1 have a synergistic effect in the development of DLBCL.
The proliferation index was correlated to the degree of the expression of HAT1 and HDAC1 in cases of DLBCL.
Following an analysis of the survival curve, the degree of the expression of HAT1 and HDAC1 was higher in association with the aggressiveness and a poor survival in cases of DLBCL. In addition, HDAC1- and HDAC2-specific inhibitors would be more effective for the treatment of DLBCL and that of PTCL-NOS and NKTCL, respectively.
- Our results indicate not only that the degree of the expression of HAT1 and HDAC1 might have a prognostic value in predicting the clinical behavior of DLBCL but also that the specific use of HDAC inhibitors might lead to better treatment outcomes of the chemotherapy in cases of malignant lymphoma.
DISCUSSION
- 1. Orr JA, Hamilton PW. Histone acetylation and chromatin pattern in cancer: a review. Anal Quant Cytol Histol 2007; 29: 17-31. PubMed
- 2. Agata Y, Katakai T, Ye SK, et al. Histone acetylation determines the developmentally regulated accessibility for T cell receptor gamma gene recombination. J Exp Med 2001; 193: 873-880. PubMedPMC
- 3. Ropero S, Esteller M. The role of histone deacetylases (HDACs) in human cancer. Mol Oncol 2007; 1: 19-25. ArticlePubMedPMCPDF
- 4. Yasui W, Oue N, Ono S, Mitani Y, Ito R, Nakayama H. Histone acetylation and gastrointestinal carcinogenesis. Ann N Y Acad Sci 2003; 983: 220-231. ArticlePubMed
- 5. Glass CK, Rosenfeld MG. The coregulator exchange in transcriptional functions of nuclear receptors. Genes Dev 2000; 14: 121-141. ArticlePubMed
- 6. Lin RJ, Sternsdorf T, Tini M, Evans RM. Transcriptional regulation in acute promyelocytic leukemia. Oncogene 2001; 20: 7204-7215. ArticlePubMedPDF
- 7. Johnstone RW. Histone-deacetylase inhibitors: novel drugs for the treatment of cancer. Nat Rev Drug Discov 2002; 1: 287-299. ArticlePubMedPDF
- 8. Dhordain P, Lin RJ, Quief S, et al. The LAZ3(BCL-6) oncoprotein recruits a SMRT/mSIN3A/histone deacetylase containing complex to mediate transcriptional repression. Nucleic Acids Res 1998; 26: 4645-4651. ArticlePubMedPMC
- 9. Peinado H, Ballestar E, Esteller M, Cano A. Snail mediates E-cadherin repression by the recruitment of the Sin3A/histone deacetylase 1 (HDAC1)/HDAC2 complex. Mol Cell Biol 2004; 24: 306-319. ArticlePubMedPMCPDF
- 10. Hajra KM, Fearon ER. Cadherin and catenin alterations in human cancer. Genes Chromosomes Cancer 2002; 34: 255-268. ArticlePubMed
- 11. Azimzadeh O, Barjaktarovic Z, Aubele M, et al. Formalin-fixed paraffin-embedded (FFPE) proteome analysis using gel-free and gel-based proteomics. J Proteome Res 2010; 9: 4710-4720. ArticlePubMed
- 12. Park Y, Kim W, Kim AY, et al. Normal prion protein in Drosophila enhances the toxicity of pathogenic polyglutamine proteins and alters susceptibility to oxidative and autophagy signaling modulators. Biochem Biophys Res Commun 2011; 404: 638-645. ArticlePubMed
- 13. Jones PA, Baylin SB. The fundamental role of epigenetic events in cancer. Nat Rev Genet 2002; 3: 415-428. ArticlePubMedPDF
- 14. Yoon SO, Kim YA, Jeon YK, Kim JE, Kang GH, Kim CW. Diffuse large B cell lymphoma shows distinct methylation profiles of the tumor suppressor genes among the non-Hodgkin's lymphomas. Korean J Pathol 2008; 42: 16-20.
- 15. Choi JH, Kwon HJ, Yoon BI, et al. Expression profile of histone deacetylase 1 in gastric cancer tissues. Jpn J Cancer Res 2001; 92: 1300-1304. ArticlePubMedPMC
- 16. Halkidou K, Gaughan L, Cook S, Leung HY, Neal DE, Robson CN. Upregulation and nuclear recruitment of HDAC1 in hormone refractory prostate cancer. Prostate 2004; 59: 177-189. ArticlePubMed
- 17. Mariadason JM. HDACs and HDAC inhibitors in colon cancer. Epigenetics 2008; 3: 28-37. ArticlePubMed
- 18. Ishihama K, Yamakawa M, Semba S, et al. Expression of HDAC1 and CBP/p300 in human colorectal carcinomas. J Clin Pathol 2007; 60: 1205-1210. ArticlePubMedPMC
- 19. Zhang Z, Yamashita H, Toyama T, et al. Quantitation of HDAC1 mRNA expression in invasive carcinoma of the breast. Breast Cancer Res Treat 2005; 94: 11-16. ArticlePubMedPDF
- 20. Huang BH, Laban M, Leung CH, et al. Inhibition of histone deacetylase 2 increases apoptosis and p21Cip1/WAF1 expression, independent of histone deacetylase 1. Cell Death Differ 2005; 12: 395-404. ArticlePubMedPDF
- 21. Song J, Noh JH, Lee JH, et al. Increased expression of histone deacetylase 2 is found in human gastric cancer. APMIS 2005; 113: 264-268. ArticlePubMed
- 22. Zhu P, Martin E, Mengwasser J, Schlag P, Janssen KP, Göttlicher M. Induction of HDAC2 expression upon loss of APC in colorectal tumorigenesis. Cancer Cell 2004; 5: 455-463. ArticlePubMed
- 23. Wilson AJ, Byun DS, Popova N, et al. Histone deacetylase 3 (HDAC3) and other class I HDACs regulate colon cell maturation and p21 expression and are deregulated in human colon cancer. J Biol Chem 2006; 281: 13548-13558. ArticlePubMed
- 24. Marquard L, Poulsen CB, Gjerdrum LM, et al. Histone deacetylase 1, 2, 6 and acetylated histone H4 in B- and T-cell lymphomas. Histopathology 2009; 54: 688-698. ArticlePubMed
- 25. Marquard L, Gjerdrum LM, Christensen IJ, Jensen PB, Sehested M, Ralfkiaer E. Prognostic significance of the therapeutic targets histone deacetylase 1, 2, 6 and acetylated histone H4 in cutaneous T-cell lymphoma. Histopathology 2008; 53: 267-277. ArticlePubMedPMC
- 26. Sugimoto T, Tomita A, Hiraga J, et al. Escape mechanisms from antibody therapy to lymphoma cells: downregulation of CD20 mRNA by recruitment of the HDAC complex and not by DNA methylation. Biochem Biophys Res Commun 2009; 390: 48-53. ArticlePubMed
- 27. Gloghini A, Buglio D, Khaskhely NM, et al. Expression of histone deacetylases in lymphoma: implication for the development of selective inhibitors. Br J Haematol 2009; 147: 515-525. ArticlePubMedPMC
REFERENCES
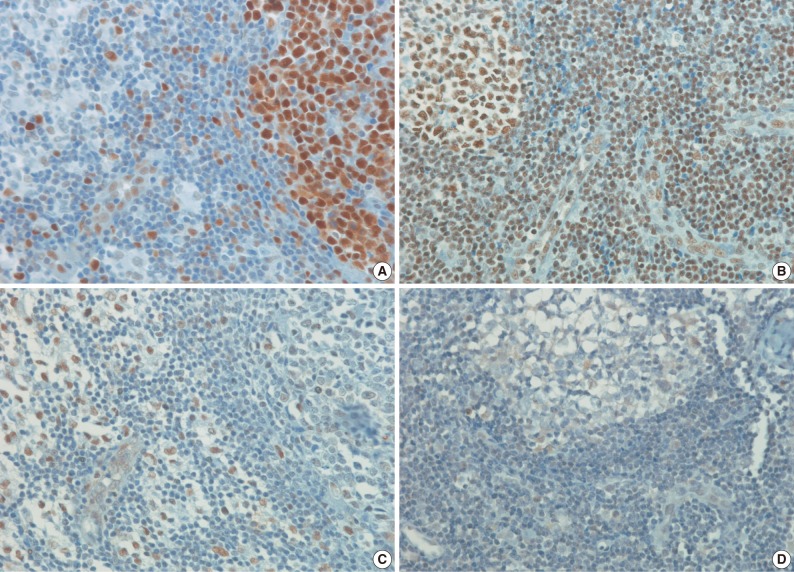
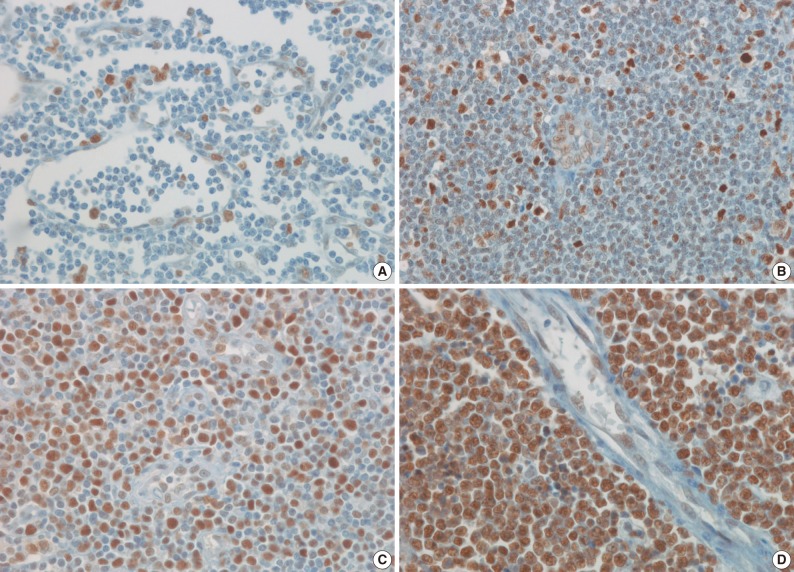
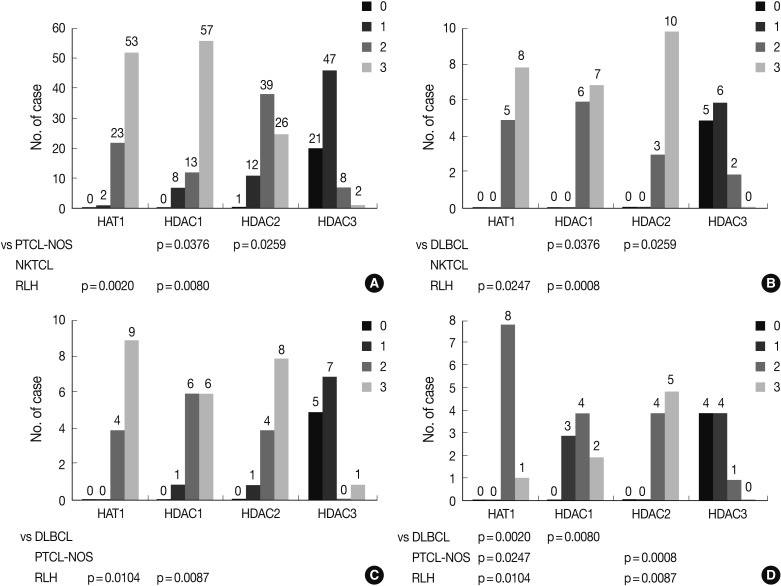
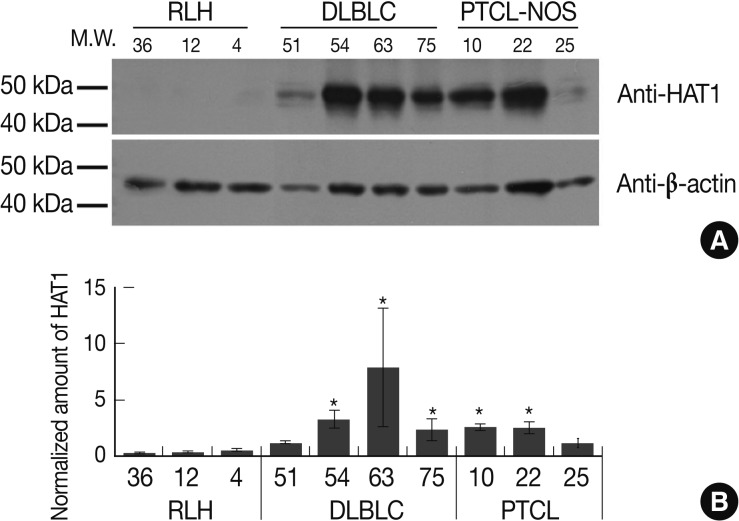
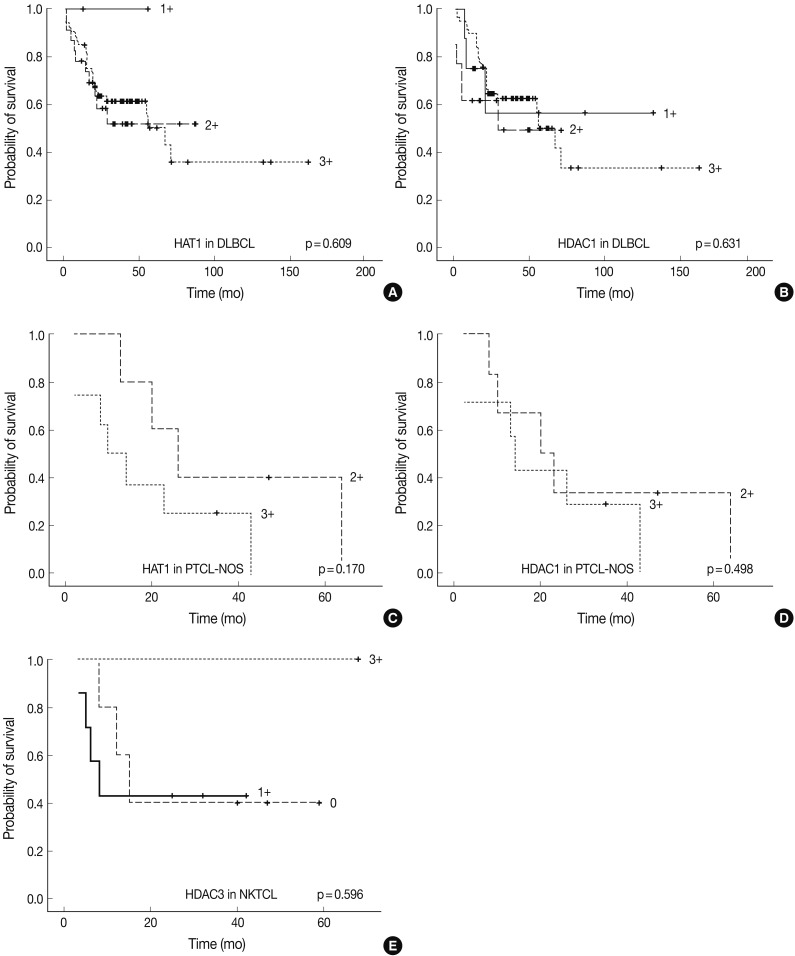
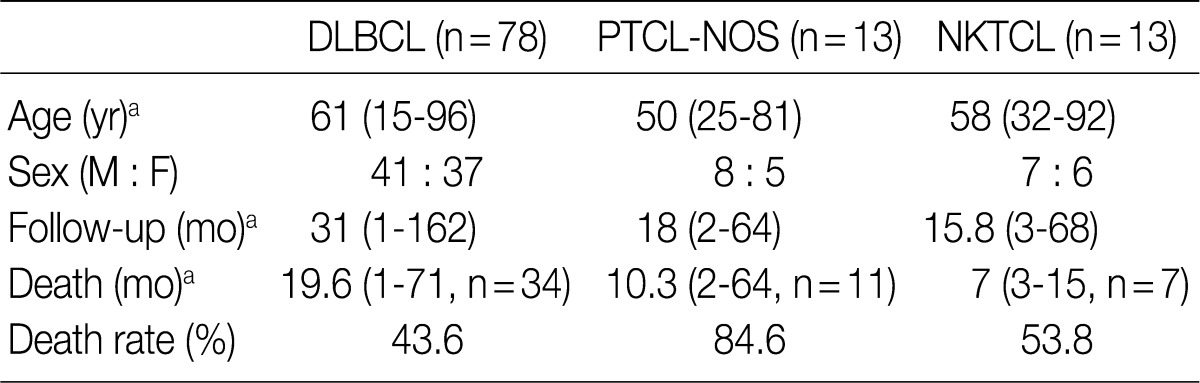
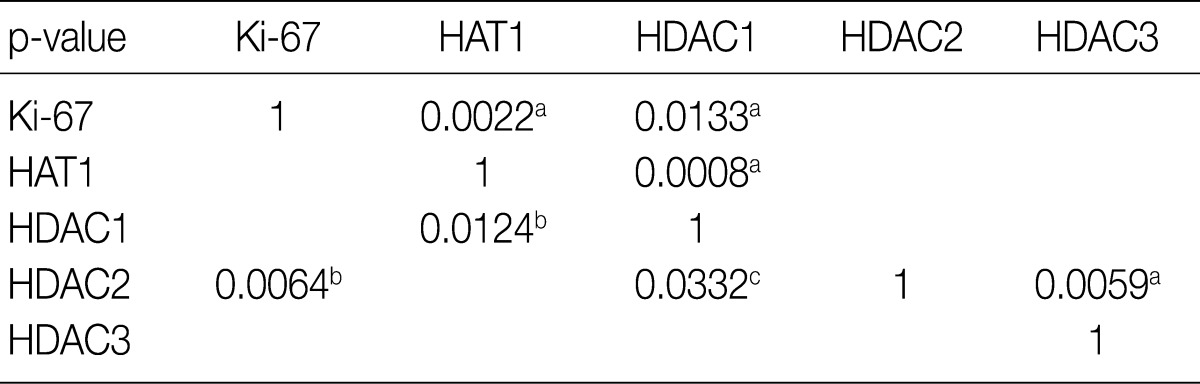
Figure & Data
References
Citations
- Low-dose chidamide maintenance therapy following allogeneic hematopoietic cell transplantation in T-cell acute lymphoblastic leukemia or lymphomas: a phase II, open-label, multicenter, single-arm trial
Yanmin Zhao, Weihao Chen, Yi Yu, Yi Luo, Jian Yu, Huarui Fu, Xiaoyu Lai, Lizhen Liu, Guifang Ouyang, Jimin Shi, Lixia Sheng, Weijie Cao, He Huang
The Lancet Regional Health - Western Pacific.2026; 72: 101916. CrossRef - Recent Advancements in the Development of HDAC/Tubulin Dual-Targeting Inhibitors
Christine Tran, Abdallah Hamze
Pharmaceuticals.2025; 18(3): 341. CrossRef - Management of T‐cell malignancies: Bench‐to‐bedside targeting of epigenetic biology
Ariana Sabzevari, Johnson Ung, Jeffrey W. Craig, Kallesh D. Jayappa, Ipsita Pal, David J. Feith, Thomas P. Loughran, Owen A. O’Connor
CA: A Cancer Journal for Clinicians.2025; 75(4): 282. CrossRef - Chidamide and orelabrutinib synergistically induce cell cycle arrest and apoptosis in diffuse large B-cell lymphoma by regulating the PI3K/AKT/mTOR pathway
Chunyan Wu, Shilv Chen, Zhimin Wu, Jiao Xue, Wen Zhang, Shan Wang, Xindong Zhao, Shaoling Wu
Journal of Cancer Research and Clinical Oncology.2024;[Epub] CrossRef - Targeting HDACs for diffuse large B-cell lymphoma therapy
Chunyan Wu, Qiao Song, Sophie Gao, Shaoling Wu
Scientific Reports.2024;[Epub] CrossRef - Advances in epigenetic therapies for B-cell non-hodgkin lymphoma
Weiwen Hu, Lanlan Zang, Xiaoxi Feng, Shuhui Zhuang, Liudi Chang, Yongjing Liu, Jinyan Huang, Yuanyuan Zhang
Annals of Hematology.2024; 103(12): 5085. CrossRef - Epigenetic regulation in hematopoiesis and its implications in the targeted therapy of hematologic malignancies
Ailin Zhao, Hui Zhou, Jinrong Yang, Meng Li, Ting Niu
Signal Transduction and Targeted Therapy.2023;[Epub] CrossRef - Understanding HAT1: A Comprehensive Review of Noncanonical Roles and Connection with Disease
Miguel A. Ortega, Diego De Leon-Oliva, Cielo Garcia-Montero, Oscar Fraile-Martinez, Diego Liviu Boaru, María del Val Toledo Lobo, Ignacio García-Tuñón, Mar Royuela, Natalio García-Honduvilla, Julia Bujan, Luis G. Guijarro, Melchor Alvarez-Mon, Miguel Ánge
Genes.2023; 14(4): 915. CrossRef - HAT1: Landscape of Biological Function and Role in Cancer
Vincenza Capone, Laura Della Torre, Daniela Carannante, Mehrad Babaei, Lucia Altucci, Rosaria Benedetti, Vincenzo Carafa
Cells.2023; 12(7): 1075. CrossRef - Recent advancement of HDAC inhibitors against breast cancer
Syed Abdulla Mehmood, Kantrol Kumar Sahu, Sounok Sengupta, Sangh Partap, Rajshekhar Karpoormath, Brajesh Kumar, Deepak Kumar
Medical Oncology.2023;[Epub] CrossRef - Noncoding rules of survival: epigenetic regulation of normal and malignant hematopoiesis
LaShanale Wallace, Esther A. Obeng
Frontiers in Molecular Biosciences.2023;[Epub] CrossRef - Potential Therapeutic Use of Aptamers against HAT1 in Lung Cancer
José Ignacio Klett-Mingo, Celia Pinto-Díez, Julio Cambronero-Plaza, Rebeca Carrión-Marchante, Miriam Barragán-Usero, María Isabel Pérez-Morgado, Eulalia Rodríguez-Martín, Mª Val Toledo-Lobo, Víctor M González, M. Elena Martín
Cancers.2022; 15(1): 227. CrossRef - Modulation of serine/threonine-protein phosphatase 1 (PP1) complexes: A promising approach in cancer treatment
Bárbara Matos, John Howl, Carmen Jerónimo, Margarida Fardilha
Drug Discovery Today.2021; 26(11): 2680. CrossRef - Histone acetyltransferase 1 promotes gemcitabine resistance by regulating the PVT1/EZH2 complex in pancreatic cancer
Yan Sun, Dianyun Ren, Yingke Zhou, Jian Shen, Heshui Wu, Xin Jin
Cell Death & Disease.2021;[Epub] CrossRef - Deciphering genes associated with diffuse large B-cell lymphoma with lymphomatous effusions: A mutational accumulation scoring approach
Sina Abdollahi, Seyedeh Zahra Dehghanian, Liang-Yi Hung, Shiang-Jie Yang, Dao-Peng Chen, L. Jeffrey Medeiros, Jung-Hsien Chiang, Kung-Chao Chang
Biomarker Research.2021;[Epub] CrossRef - The contributory roles of histone deacetylases (HDACs) in hematopoiesis regulation and possibilities for pharmacologic interventions in hematologic malignancies
Mahdieh Mehrpouri, Atieh Pourbagheri-Sigaroodi, Davood Bashash
International Immunopharmacology.2021; 100: 108114. CrossRef - Emerging role of histone deacetylase inhibitors in the treatment of diffuse large B-cell lymphoma
Mingyang Wang, Xiaosheng Fang, Xin Wang
Leukemia & Lymphoma.2020; 61(4): 763. CrossRef Effective Treatment with PD-1 Antibody, Chidamide, Etoposide, and Thalidomide (PCET) for Relapsed/Refractory Natural Killer/T-Cell Lymphoma: A Report of Three Cases
Lijun Du, Lei Zhang, Ling Li, Xin Li, Jiaqin Yan, Xinhua Wang, Xiaorui Fu, Zhenchang Sun, Xudong Zhang, Zhaoming Li, Jingjing Wu, Hui Yu, Yu Chang, Zhiyuan Zhou, Feifei Nan, Xiaolong Wu, Li Tian, Mingzhi Zhang
OncoTargets and Therapy.2020; Volume 13: 7189. CrossRef- Overexpressed histone acetyltransferase 1 regulates cancer immunity by increasing programmed death-ligand 1 expression in pancreatic cancer
Ping Fan, Jingyuan Zhao, Zibo Meng, Heyu Wu, Bo Wang, Heshui Wu, Xin Jin
Journal of Experimental & Clinical Cancer Research.2019;[Epub] CrossRef - Histone modifications: A review about the presence of this epigenetic phenomenon in carcinogenesis
Emanuely Silva Chrun, Filipe Modolo, Filipe Ivan Daniel
Pathology - Research and Practice.2017; 213(11): 1329. CrossRef - Histone Acetyltransferase 1 Promotes Cell Proliferation and Induces Cisplatin Resistance in Hepatocellular Carcinoma
Xin Jin, Shenghua Tian, Pingping Li
Oncology Research Featuring Preclinical and Clinical Cancer Therapeutics.2017; 25(6): 939. CrossRef - HDACs and HDAC Inhibitors in Cancer Development and Therapy
Yixuan Li, Edward Seto
Cold Spring Harbor Perspectives in Medicine.2016; 6(10): a026831. CrossRef - Histone deacetylase inhibitors and epigenetic regulation in lymphoid malignancies
Diana Markozashvili, Vincent Ribrag, Yegor S. Vassetzky
Investigational New Drugs.2015; 33(6): 1280. CrossRef - Genome-Wide Association Study of Event-Free Survival in Diffuse Large B-Cell Lymphoma Treated With Immunochemotherapy
Hervé Ghesquieres, Susan L. Slager, Fabrice Jardin, Amelie S. Veron, Yan W. Asmann, Matthew J. Maurer, Thierry Fest, Thomas M. Habermann, Marie C. Bene, Anne J. Novak, Sylvain Mareschal, Corinne Haioun, Thierry Lamy, Stephen M. Ansell, Herve Tilly, Thomas
Journal of Clinical Oncology.2015; 33(33): 3930. CrossRef - Histone deacetylase 2 controls p53 and is a critical factor in tumorigenesis
Tobias Wagner, Peter Brand, Thorsten Heinzel, Oliver H. Krämer
Biochimica et Biophysica Acta (BBA) - Reviews on Cancer.2014; 1846(2): 524. CrossRef - Targetome profiling and functional genetics implicate miR-618 in lymphomagenesis
Alan Fu, Aaron E Hoffman, Ran Liu, Daniel I Jacobs, Tongzhang Zheng, Yong Zhu
Epigenetics.2014; 9(5): 730. CrossRef - Expression of Histone Deacetylases HDAC1, HDAC2, HDAC3, and HDAC6 in Invasive Ductal Carcinomas of the Breast
Jinwon Seo, Soo Kee Min, Hye-Rim Park, Dong Hoon Kim, Mi Jung Kwon, Lee Su Kim, Young-Su Ju
Journal of Breast Cancer.2014; 17(4): 323. CrossRef - Diffuse large B-cell lymphoma
Maurizio Martelli, Andrés J.M. Ferreri, Claudio Agostinelli, Alice Di Rocco, Michael Pfreundschuh, Stefano A. Pileri
Critical Reviews in Oncology/Hematology.2013; 87(2): 146. CrossRef - Histone deacetylase inhibitors activate CIITA and MHC class II antigen expression in diffuse large B‐cell lymphoma
Kelly A. Cycon, Kathleen Mulvaney, Lisa M. Rimsza, Daniel Persky, Shawn P. Murphy
Immunology.2013; 140(2): 259. CrossRef
 PubReader
PubReader Cite this Article
Cite this Article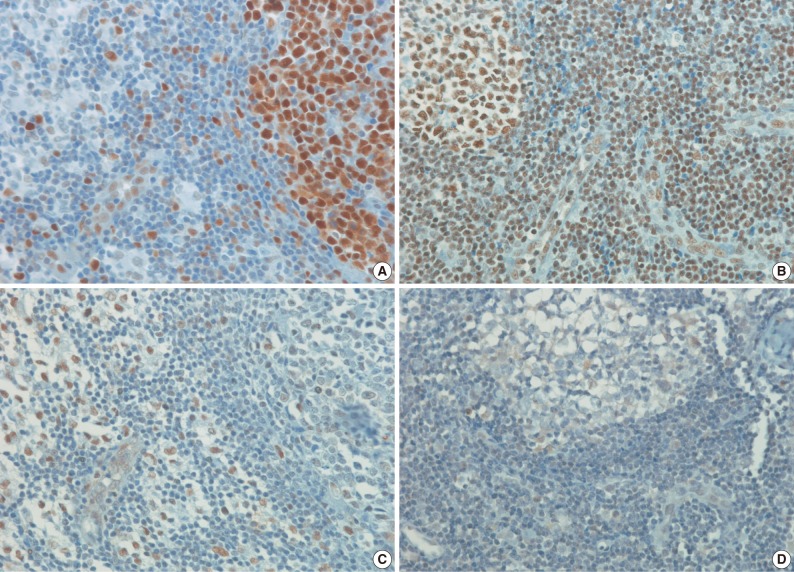
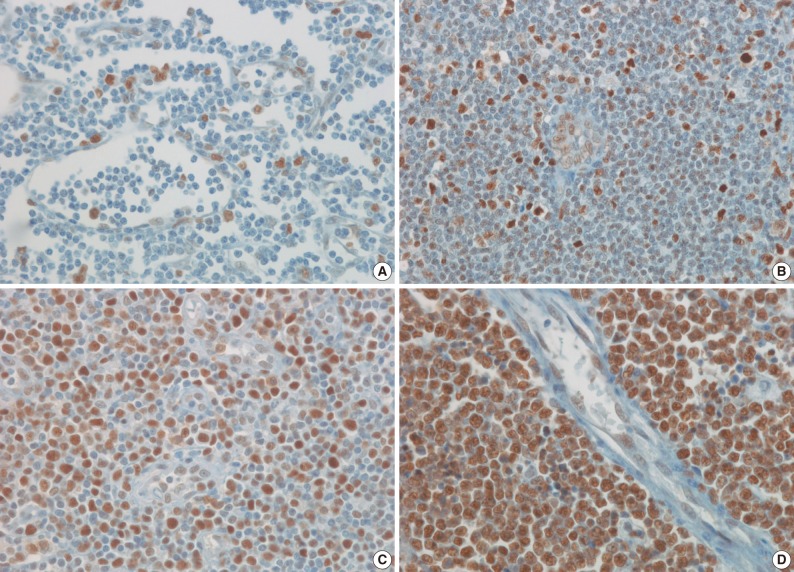
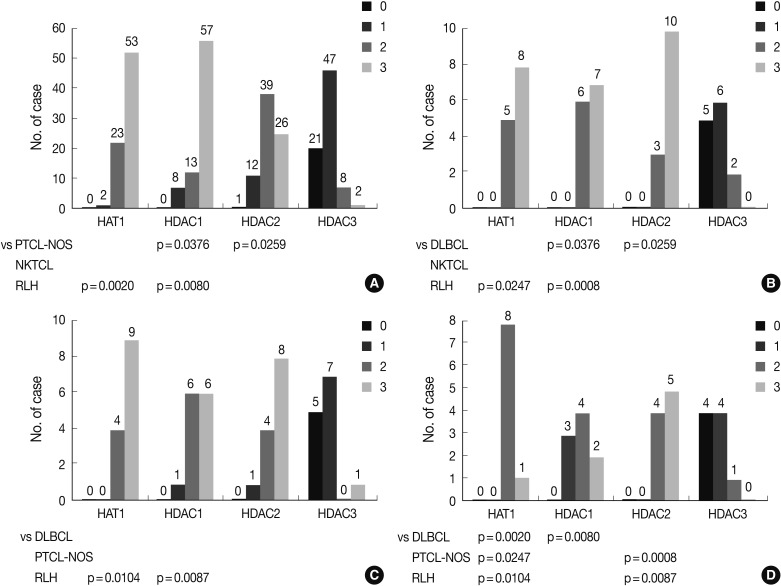
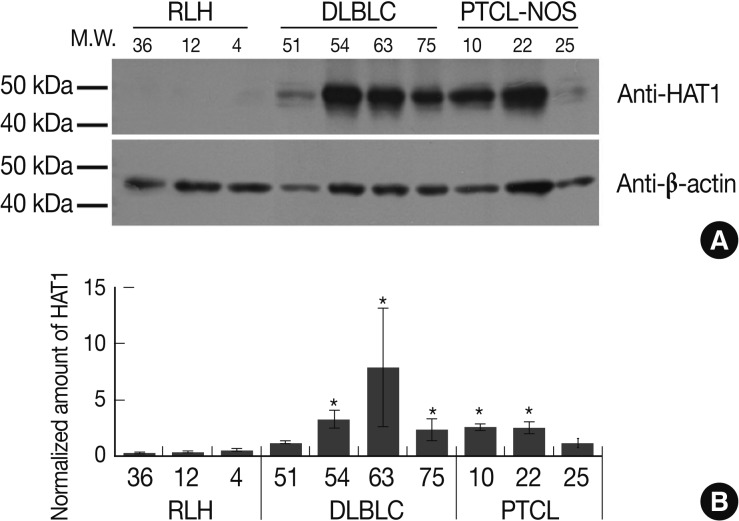





Fig. 1
Fig. 2
Fig. 3
Fig. 4
Fig. 5

IHCs, immunohistochemical staining; HAT1, histone acetyltransferase 1; HDAC, histone deacetylase.
DLBCL, diffuse large B-cell lymphomas; PTCL-NOS, peripheral T-cell lymphomas, not otherwise specified; NKTCL, NK/T-cell lymphomas, nasal type; M, male; F, female. aValues are presented as mean (range).
HAT1, histone acetyltransferase 1; HDAC, histone deacetylase. Blank, p>0.05 in three types of lymphomas; aDiffuse large B-cell lymphomas; bNK/T-cell lymphomas, nasal type; cPeripheral T-cell lymphomas, not otherwise specified.

