PLUNC downregulates the expression of PD-L1 by inhibiting the interaction of DDX17/β-catenin in nasopharyngeal carcinoma
Article information

Abstract
Background
Nasopharyngeal carcinoma (NPC) is characterized by high programmed death-ligand 1 (PD-L1) expression and abundant infiltration of non-malignant lymphocytes, which renders patients potentially suitable candidates for immune checkpoint blockade therapies. Palate, lung, and nasal epithelium clone (PLUNC) inhibit the growth of NPC cells and enhance cellular apoptosis and differentiation. Currently, the relationship between PLUNC (as a tumor-suppressor) and PD-L1 in NPC is unclear.
Methods
We collected clinical samples of NPC to verify the relationship between PLUNC and PD-L1. PLUNC plasmid was transfected into NPC cells, and the variation of PD-L1 was verified by western blot and immunofluorescence. In NPC cells, we verified the relationship of PD-L1, activating transcription factor 3 (ATF3), and β-catenin by western blot and immunofluorescence. Later, we further verified that PLUNC regulates PD-L1 through β-catenin. Finally, the effect of PLUNC on β-catenin was verified by co-immunoprecipitation (Co-IP).
Results
We found that PLUNC expression was lower in NPC tissues than in paracancer tissues. PD-L1 expression was opposite to that of PLUNC. Western blot and immunofluorescence showed that β-catenin could upregulate ATF3 and PD-L1, while PLUNC could downregulate ATF3/PD-L1 by inhibiting the expression of β-catenin. PLUNC inhibits the entry of β-catenin into the nucleus. Co-IP experiments demonstrated that PLUNC inhibited the interaction of DEAD-box helicase 17 (DDX17) and β-catenin.
Conclusions
PLUNC downregulates the expression of PD-L1 by inhibiting the interaction of DDX17/β-catenin in NPC.
INTRODUCTION
The 2020 Global Cancer Statistics report showed that there were more than 133,000 new cases of nasopharyngeal cancer (NPC) worldwide [1]. NPC occurs mainly in East and South Asia. NPC is sensitive to radiotherapy, so radiotherapy is the preferred treatment. Radiotherapy plus adjuvant chemotherapy is the standard treatment for advanced NPC. However, 20%–30% of NPC patients will experience recurrence, with most recurrences occurring within the first 2 years after treatment [2]. Because of its unique immune environment, NPC is considered a highly immune-inflamed tumor. Massive lymphocytic infiltrations, high programmed death-ligand 1 (PD-L1) expression, and several key immune molecules that regulate the activation of T-cells (CD40, CD70, CD80, and CD86) are often observed in Epstein-Barr virus (EBV)–induced NPC [3]. Since NPC is characterized by high PD-L1 expression and significant non-malignant lymphocyte infiltration, patients may be suitable for immune checkpoint-blocking therapies [4]. Fortunately, anti–programmed death-1 (PD-1)/PD-L1 treatment has made certain progress in metastatic or recurrent metastatic NPC as a promising method to prolong the survival rate of patients [5]. Therefore, understanding the regulatory mechanism of PD-L1 is of great significance for the immunotherapy of NPC.
Palatal, lung, and nasal epithelial clone (PLUNC) protein is encoded by a 300-KB gene segment on chromosome 20, which is specifically expressed in the upper airway and nasopharyngeal regions and is involved in host defense [6,7]. PLUNC contributes to the overall surface tension of airway epithelial secretions, and this activity may interfere with the formation of biofilms in airway pathogens [8,9]. In innate immune defense, the PLUNC protein is involved in antimicrobial activity and can directly kill Escherichia coli and Acinetobacter haemolyticus through cell wall permeability [10]. In patients with chronic sinusitis, reduced levels of PLUNC expression are associated with repeated sinus surgery and the amount of bacterial colonization [11]. In acute pulmonary exacerbations (AEs), PLUNC expression decreases sharply as inflammation increases. In stable patients, lower PLUNC expression correlates with an increased AE risk. Thus, the downregulation of PLUNC could predict AE at the early stages [12]. However, in gastric hepatoid adenocarcinoma, PLUNC expression may be an important factor that can indicate the tumor's malignant potential, as positive cases show vascular invasion and lymph node metastasis [13].
A previous study suggested that PLUNC was downregulated in NPC and related to the poor prognosis of NPC patients [14]. PLUNC can inhibit the tumor inflammatory microenvironment by regulating the Toll-like receptor 9/nuclear factor κB signaling pathway, thereby reducing the inflammatory response of EBV-induced NPC cells [15]. PLUNC may be a tumor-suppressor gene in NPC; in previous research, it inhibited the growth of NPC cells and promoted cellular apoptosis and differentiation [16,17]. Our results revealed that PD-L1 is highly expressed in nasopharyngeal carcinoma. However, the relationship between PLUNC and PD-L1 in NPC is unclear.
Wnt/β-catenin signaling plays a crucial role in cancer. Abnormal Wnt/β-catenin signaling had been found to be closely related to the occurrence, development, and malignant transformation of cancer [18]. β-catenin is highly expressed in NPC tissues, and its positive expression negatively correlates with the survival rate of NPC patients [19]. It had been reported that β-catenin promotes the proliferation and metastasis of NPC cells [20,21]. However, it is still unclear whether β-catenin regulates PD-L1 in NPC.
In our study, we found a negative correlation between PLUNC and PD-L1 expression in NPC, and PLUNC suppressed PD-L1 expression by downregulating β-catenin. Mechanically, PLUNC downregulated PD-L1 expression by inhibiting DEAD-box helicase 17 (DDX17)/β-catenin interaction. In summary, our research will provide new ideas for immune checkpoint blockade therapy in NPC.
MATERIALS AND METHODS
Bioinformatic analysis
We conducted gene expression differential analysis using a t test based on data from head and neck cancer cases in the GEPIA public database(http://gepia.cancer-pku.cn/detail.php. In addition, the differences in EBV infection between high- and low-expression groups of the PLUNC gene in the GSE102349 dataset from the Gene Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo/GEO) were compared using the t test, and the correlation between EBV infection and PLUNC gene expression was analyzed using Pearson’s correlation analysis.
Cell culture
NPC cell lines (5-8F, HNE2, S18, and HONE1) and the 293T cell line come from the Cancer Research Institute of Central South University. 5-8F, HNE2, S18, and HONE1 were used for the study of protein expression, localization, and signal transduction in nasopharyngeal carcinoma cells. 293T cells are the most commonly used cells in the study of protein interactions in articles. These cells were maintained at 37°C and 5% CO2 in Dulbecco’s modified Eagle medium supplemented with 10% fetal bovine serum (Biological Industries, Kibbutz Beit-Haemek, Israel), 100 U/mL of penicillin, and 100 μg/mL of streptomycin (GE Healthcare, Waukesha, WI, USA).
Cell transfection
Transfection plasmids (including PLUNC overexpression), PLUNC control vectors, PLUNC/β-catenin/activating transcription factor 3 (ATF3) small hairpin RNAs (shRNAs), and their negative controls were purchased from Shanghai Genechem (Shanghai, China). DDX17/β-catenin/ATF3 lentiviral expression vectors were constructed by inserting expanded DDX17/β-catenin/ATF3 cDNA fragments into a lentiviral shuttle vector. The cells were seeded into six-well plates, and, upon reaching 80% confluence, recombinant plasmids were transfected using Lipofectamine 2000 (Thermo Fisher Scientific, Waltham, MA, USA), following the manufacturer's protocol. The indicated cells infected with the recombinant lentiviral vectors were selected with puromycin or G418 for 2 weeks.
Co-immunoprecipitation assay
Cell lysis ice immunoprecipitation lysis buffer was supplemented with protease and phosphatase inhibitors for 15 minutes. Add immunoprecipitation lysis buffer containing protease inhibitors and phosphatase inhibitors to the cells and lyse on ice for 15 minutes. After cell lysis, centrifuge at 16,000 ×g for 10 minutes at 4°C and incubate the supernatant with antibodies. Following incubation, the cells were added into pre-washed agarose beads, coupled with protein G or protein A, and incubated at 4°C for 3 hours. The immune complex was purified on the magnet by extensive washing of the lysis buffer. The chemiluminescence signals were collected by a BIO-RAD Gel imaging analysis system scanner (Bio-Rad Laboratories, Hercules, CA, USA).
Western blot analysis
Cells were lysed with RIPA lysis buffer, and the protein concentration was measured using a bicinchoninic acid protein assay kit (KeyGen Biotech, Nanjing, China). Protein samples (30 μg/well) were loaded onto 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and then transferred onto polyvinylidene difluoride membranes. After blocking with skim milk, membranes were incubated in primary antibodies at 4°C overnight and then in horseradish peroxidase–conjugated secondary antibodies for 1 hour at 37°C. Primary antibodies used for experiments included anti-PLUNC (ab131163, Abcam, Cambridge, UK), glyceraldehyde 3-phosphate dehydrogenase (Abcam), anti–β-catenin, T-cell factor 4 (TCF4) (Cell Signaling Technology, Danvers, MA, USA), DDX17 (sc398168, Santa Cruz Biotechnology, Dallas, TX, USA), PD-L1 (28076-1-AP, Proteintech, Rosemont, IL, USA), ATF3 (ab254268, Abcam), glycogen synthase kinase 3β (GSK-3β; 67329-1-Ig, Proteintech), and p-GSK-3β (67558-1-Ig, Proteintech). Protein bands were imaged and captured using ECL reagent (Biosharp, Hefei City, China).
Immunofluorescence staining
Cells were seeded on coverslips, fixed with 4% paraformaldehyde, and then permeabilized with 0.2% Triton X-100. After blocking with 5% bovine serum albumin, the cells were incubated with a primary antibody (anti-ATF3 [Abcam]; anti-Plunc, anti–β-catenin, and anti–PD-L1 [Proteintech]; and anti-DDX17 [Santa Cruz Biotechnology]) overnight at 4°C, which was followed by incubation with an Alexa Fluor 488– or Alexa Fluor 594–conjugated secondary antibody (Abcam). DAPI was used to stain cell nuclei. Immunofluorescence images were captured with a fluorescence microscope (Olympus Corp., Tokyo, Japan).
Clinical NPC samples and immunohistochemistry
NPC samples and paired paracancer tissues were collected from 30 NPC patients at the Second Xiangya Hospital of Central South University (Changsha, China). The study was approved by the Joint Ethics Committee of the Central South University Health Authority, and informed consent was obtained from each participant.
For immunohistochemistry, tissue sections of formalin-fixed and paraffin-embedded NPC tissues were incubated with anti-PLUNC (10413-1-AP, Proteintech), β-catenin (51067-2-AP, Proteintech), PD-L1 (28076-1-AP, Proteintech), DDX17 (sc398168, Santa Cruz Biotechnology) or control IgG1 (1 μg/mL). After washing with phosphate buffered saline, the slides were reacted with the Prolink-2 Plus horseradish peroxidase rabbit polymer detection kit (Golden Bridge International, Bothell, WA, USA), and the images were scanned using Aperio ScanScope CS software (Aperio Technologies, Vista, CA, USA). Two independent pathologists evaluated and scored the slides based on the intensity and extent of staining (double-blinded). A staining index (values, 0–9) was obtained from the intensity of the positive staining (negative = 0 points, weak = 1 points, moderate = 2 points, strong = 3 points), and the proportion of positive cells of interest (<25% = 1 point, 25%–50% = 2 points, ≥50% = 3 points) was calculated. The final results were then designated and rescored as follows: 0 points = negative, 1–3 points = weak positive (1 point), 4–6 points = moderately positive (2 points), or 7–9 points = strongly positive (3 points). All sections were scored independently by two pathologists blinded to the clinicopathological features and the clinical course.
Statistical analysis
Data were analyzed using GraphPad Prism (GraphPad Software, La Jolla, CA, USA). Graphed data were presented as mean ± standard error of the mean values, and statistical significance was determined using Student’s t test. Pearson’s chi-squared or Fisher's exact test was used to analyze clinicopathological parameters. Multivariable Cox proportional hazards modeling was performed to identify independent risk factors for the prognosis of NPC. p ≤ 0.05 was considered statistically significant.
RESULTS
PLUNC negatively correlates with PD-L1 expression in NPC
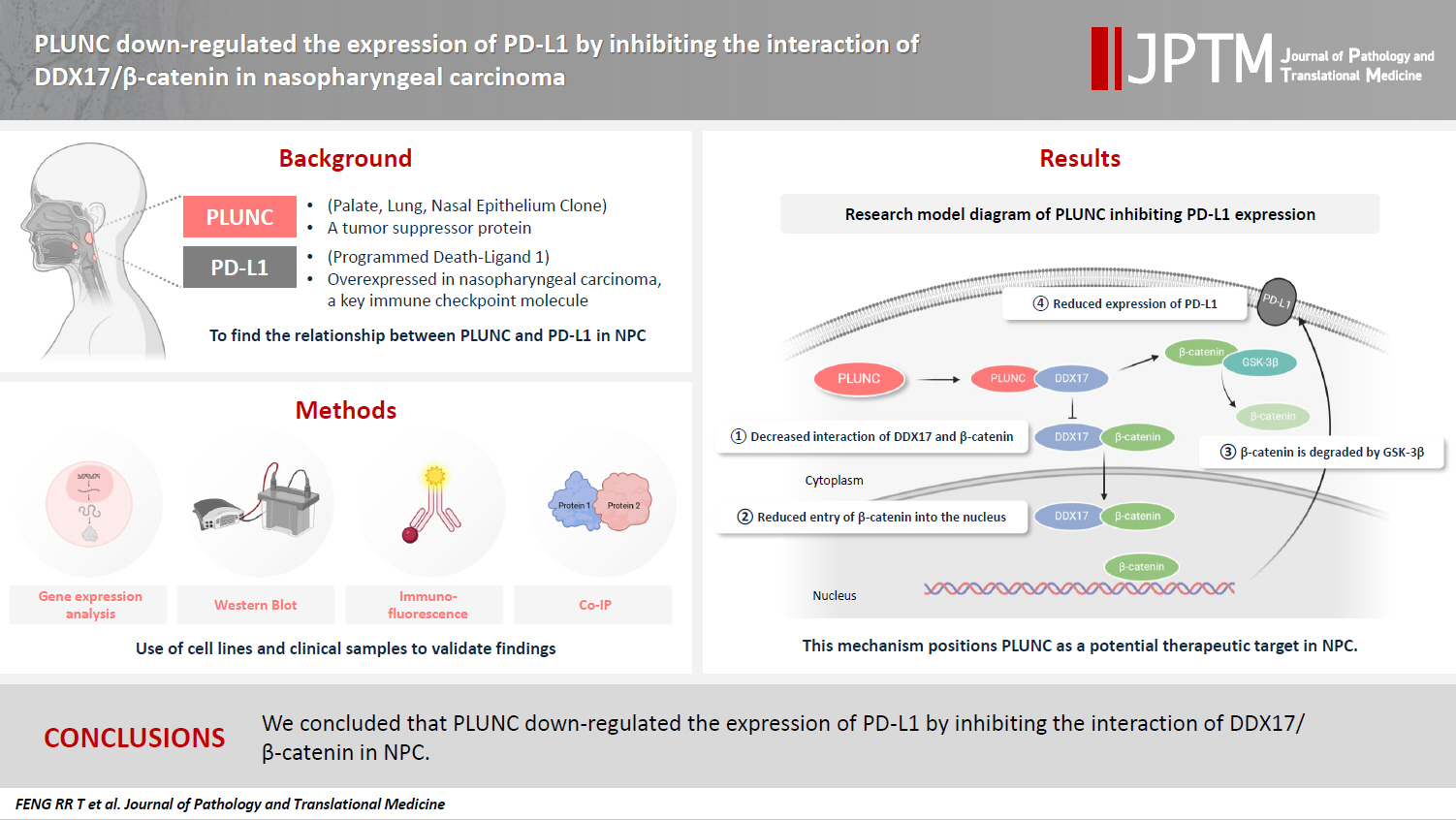
In the GEPIA online database (http://gepia.cancer-pku.cn/detail.php), the expression of the PLUNC gene (BPIFA1) was low in head and neck squamous cell carcinoma, while that of the PD-L1 gene (CD274) was high (Fig. 1A). The X-axis represents the tumor group and the normal group, while the Y-axis uses log2 (TPM+1) values to represent gene-expression levels. Based on the information of 114 NPC patients represented in tissue microarrays (Superbiotek, Shanghai, China) (for relevant clinical information, see Supplementary Table S1), we conducted univariate logistic regression analysis according to the immunohistochemical staining score of PD-L1 (0–3 points; 0 points = negative, >0 points = positive) and found that PD-L1 is a risk factor affecting the prognosis of nasopharyngeal carcinoma patients (Fig. 1B, C). Considering the close correlation between NPC occurrence and EBV infection, we analyzed the effect of EBV on PLUNC gene expression. The results showed no difference in the degree of EBV infection in NPC with high and low expression of PLUNC (Supplementary Fig. S1A). Moreover, there was no correlation between EBV infection and PLUNC gene-expression level (R = −0.16, p = 0.083) (Supplementary Fig. S1B). To investigate the correlation between PLUNC and PD-L1, we collected paraffin sections from 30 NPC patients in the Second Xiangya Hospital of Central South University for immunohistochemical staining. The results showed that the expression of PLUNC was low in NPC and high in paracancer tissues, while the expression of PD-L1 was opposite to that of PLUNC (Fig. 1D). The statistical analysis of PD-L1 scores in tumor tissues and adjacent tissues of 30 NPC patients is presented in Supplementary Fig. S1C, indicating that the expression of PD-L1 was significantly greater in tumor tissues than in paracancer tissues. The negative and positive control results of immunohistochemical staining are shown in Supplementary Fig. S1D. Next, we validated the relationship between PLUNC and PD-L1 in NPC cells. In the HNE2 and 5-8F cell lines, we overexpressed PLUNC and found that PD-L1 was downregulated (Fig. 1E). However, when PLUNC was knocked down, the expression of PD-L1 was upregulated (Fig. 1F). Therefore, we considered a negative correlation to exist between the expression of PLUNC and PD-L1 in NPC.
Palatal, lung, and nasal epithelial clone (PLUNC) negatively correlates with programmed death-ligand 1 (PD-L1) expression in nasopharyngeal carcinoma (NPC). (A) PLUNC and PD-L1 gene expressions in the head and neck squamous cell carcinoma database. T represents tumor and N represents normal. (B) The forest map of risk factor analysis for NPC tissue chips, *p < .05, ***p < .001, with significant differences. (C) Detection of PD-L1 expression levels in NPC tissue and paracancer tissues using tissue chips. (D) Immunohistochemical staining was performed on paraffin sections of NPC and adjacent tissues to detect the expression levels of PLUNC and PD-L1. (E) Verification of PD-L1 expression in 5-8F and HNE2 cell lines after overexpression of PLUNC through western blot. (F) After transfection of PLUNC-shRNA in 5-8F and HNE2 cells, western blot was used to detect the expression of PD-L1. NC, negative control; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; WBC, white blood cell; AIC, Akaike information criterion.
β-catenin upregulates ATF3/PD-L1 expression
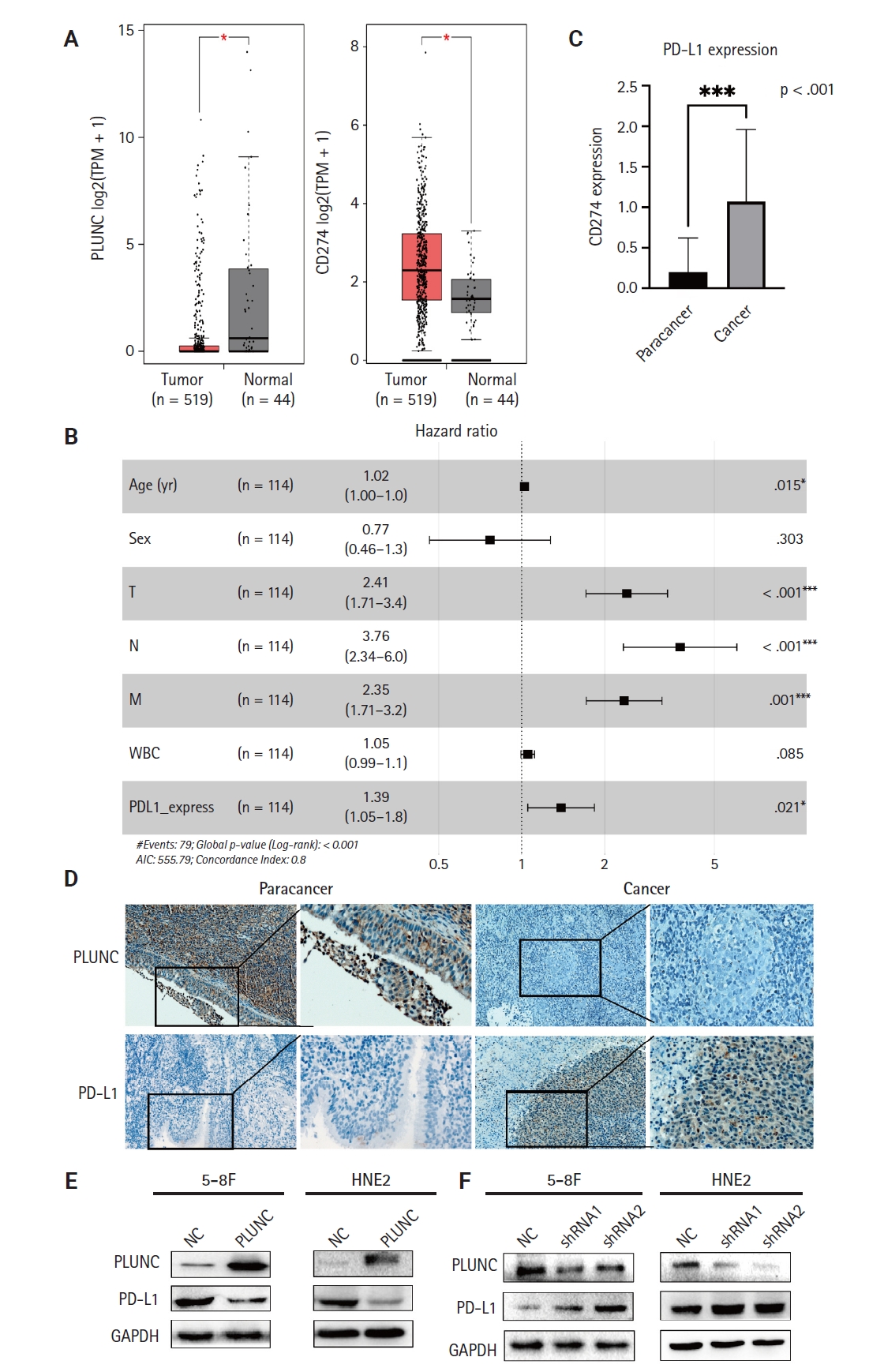
ATF3 is a transcription factor; as a member of the ATF/CREB family, it can regulate the expression of PD-L1 in melanoma cells [22]. We validated the regulation of PD-L1 by ATF3 in NPC cells. When we overexpressed ATF3 in 5-8F and S18 cells, the expression of PD-L1 was upregulated (Supplementary Fig. S2A, B). Conversely, when we knocked down ATF3, the expression of PD-L1 was downregulated (Supplementary Fig. S2C, D). The high expression of β-catenin is a risk factor for poor prognosis in NPC patients [23]. β-catenin is an upstream molecule of PD-L1 in glioblastoma [24]. However, whether β-catenin regulates PD-L1 in NPC remains unclear, so we validated the relationship between β-catenin and PD-L1 in NPC cells. When we overexpressed β-catenin in S18 cells and HNE2 cells, the expressions of ATF3 and PD-L1 were upregulated (Fig. 2A). Conversely, when we knocked down β-catenin in 5-8F and HONE1 cells, the expression of both PD-L1 and ATF3 was downregulated (Fig. 2B; statistical results are shown in Fig. 2C, D). Meanwhile, we validated the mRNA- and protein-expression levels of β-catenin after transfection with shRNA (Supplementary Fig. S3A, B). In addition, we also confirmed the expression of both ATF3 and PD-L1 through immunofluorescence experiments (Fig. 2E-H). Therefore, we confirmed that β-catenin upregulated the expression of PD-L1 and ATF3 in NPC cells.
β-catenin upregulates activating transcription factor 3 (ATF3)/programmed death-ligand 1 (PD-L1) expression. (A) Verification of PD-L1 and ATF3 expression in S18 and HNE2 cell lines by western blot after overexpression of β-catenin. (B) Verification of PD-L1 and ATF3 expression in 5-8F and HONE1 cell lines by western blot after transfection with β-catenin–shRNA. (C, D) Semi-quantitative analysis of the protein expressions of β-catenin, PD-L1, and ATF3 after overexpression and knockdown with β-catenin. *p < .05. (E, F) After overexpressing β-catenin, the expression of PD-L1 was verified by immunofluorescence. (G, H) After knocking down β-catenin, the expression of PD-L1 was verified by immunofluorescence. NC, negative control; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
PLUNC suppresses the β-catenin pathway
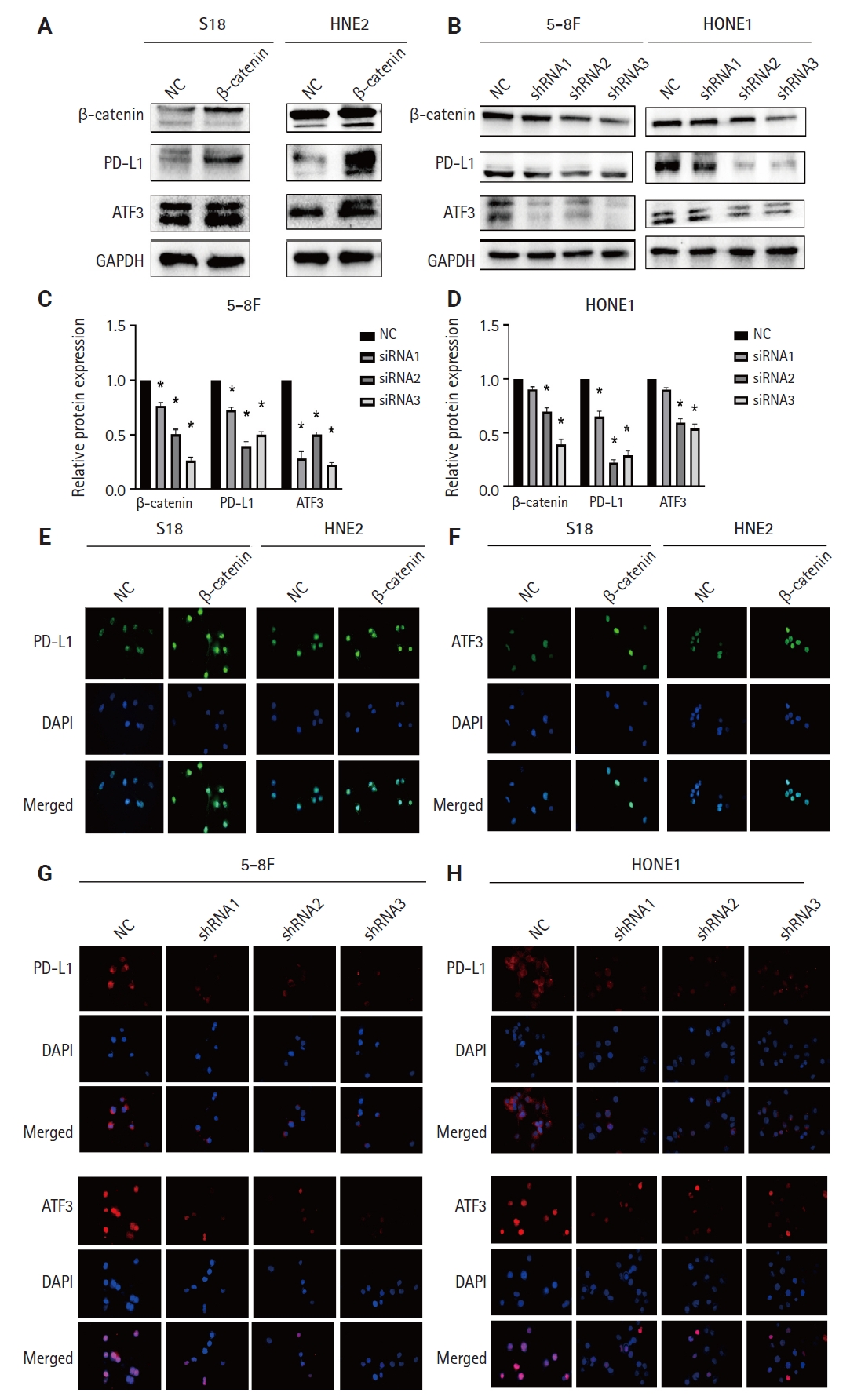
Since β-catenin could upregulate PD-L1 expression and PLUNC could downregulate PD-L1 expression, could PLUNC inhibit the β-catenin pathway? After overexpressing PLUNC in 5-8F and HNE2 cells (Supplementary Fig. S3C), we found that GSK-3β (which promotes β-catenin ubiquitination degradation) [25] was upregulated. Conversely, phosphorylated GSK-3β (its inactivated form) was upregulated. The expression of TCF4, which could interact with β-catenin to regulate downstream gene transcription [26], was also inhibited (Fig. 3A). The relative changes in β-catenin, GSK-3β, p–GSK-3β, TCF4, and ATF3 are shown in Fig. 3B and C. We also found that overexpression of PLUNC resulted in the downregulation of β-catenin expression by immunofluorescence experiments (Fig. 3D). On the contrary, knocking down PLUNC in HNE2 and 5-8F cells (Supplementary Fig. S3D) weakened the inhibition of the β-catenin pathway (Fig. 3E, F). Therefore, we considered that PLUNC could inhibit the β-catenin pathway.
Palatal, lung, and nasal epithelial clone (PLUNC) suppresses the β-catenin pathway. (A) Detection of the β-catenin pathway expression in 5-8F and HNE2 cell lines by western blot after overexpression of PLUNC. (B, C) Semi-quantitative analysis of the protein expressions of β-catenin, glycogen synthase kinase 3β (GSK-3β), p–GSK-3β, T-cell factor 4 (TCF4), and activating transcription factor 3 (ATF3) after overexpression with PLUNC. *p < .05. (D) β-catenin expression in 5-8F and HNE2 cell lines by immunofluorescence after overexpression of PLUNC. (E) Detection of β-catenin pathway expression in 5-8F and HNE2 cell lines by western blot, and the semi-quantitative analysis of the protein expression after knocking down PLUNC. (F) β-catenin expression in 5-8F and HNE2 cell lines by immunofluorescence after knocking down PLUNC. NC, negative control; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
PLUNC downregulates the expression of ATF3/PD-L1 through inhibiting the β-catenin pathway
After overexpressing PLUNC in 5-8F and HNE2 cells, we observed that the expression of ATF3/PD-L1 decreased (Fig. 4A–D). Conversely, when we knocked down PLUNC in 5-8F and HNE2 cells, we observed that the expression of both ATF3 and PD-L1 increased (Fig. 4E–H). After treating cells using XAV-939 as an inhibitor of the Wnt/β-catenin pathway, the expression of β-catenin, p-GSK3β, ATF3, and PD-L1 decreased. However, when PLUNC was knocked down in 5-8F and HNE2 cells, the expressions of β-catenin, p-GSK3β, ATF3, and PD-L1 increased. When PLUNC-shRNA was combined with XAV-939, the expressions of β-catenin, p-GSK3β, ATF3, and PD-L1 were weakened (Fig. 4I, J). Thus, we believed that PLUNC downregulated the expression of ATF3/PD-L1 through inhibiting the β-catenin pathway.
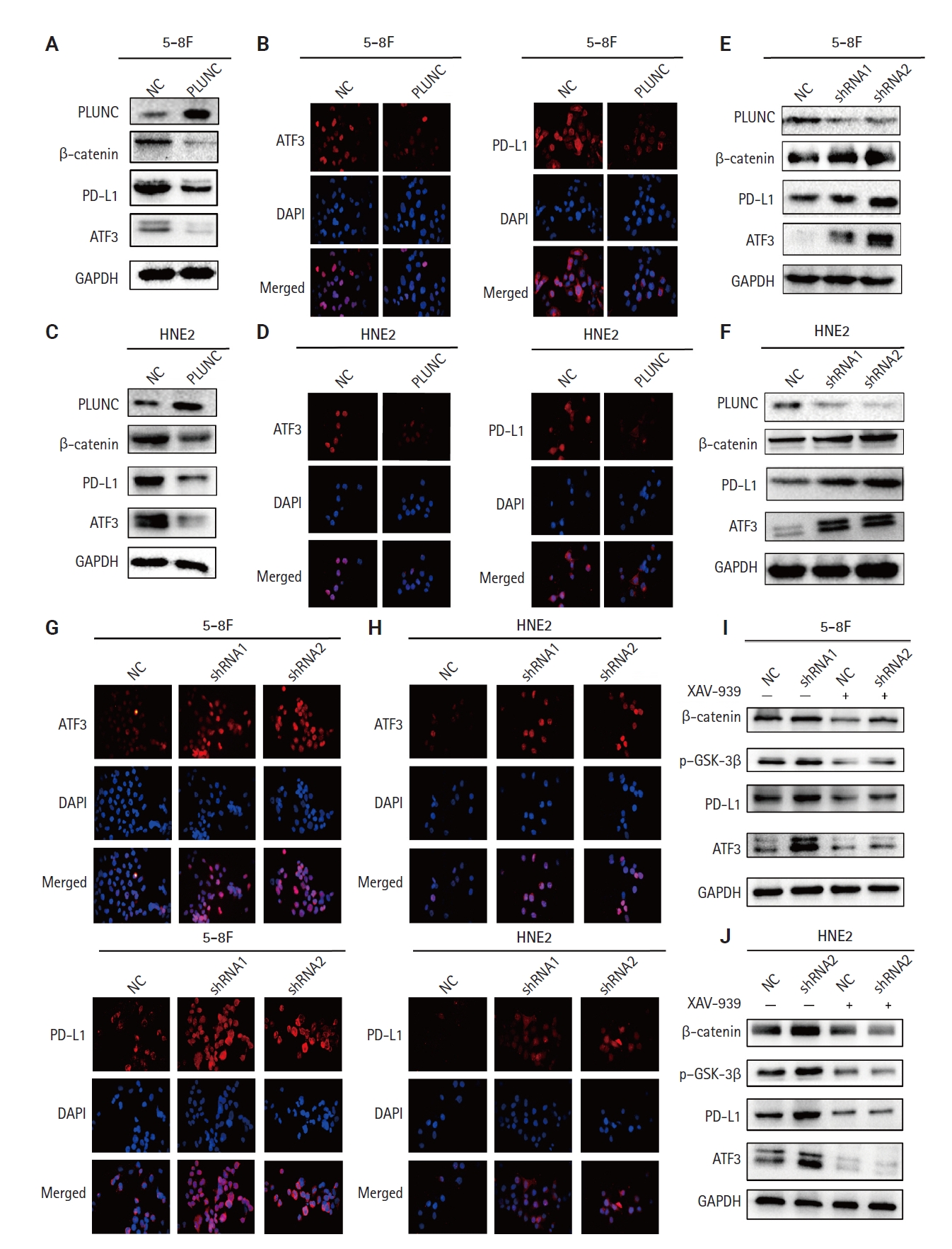
Palatal, lung, and nasal epithelial clone (PLUNC) downregulates the expression of activating transcription factor 3 (ATF3)/programmed death-ligand 1 (PD-L1) by inhibiting the β-catenin pathway. (A, B) After overexpression of PLUNC in 5-8F cells, decreased ATF3 and PD-L1 expression was confirmed by western blot and immunofluorescence. (C, D) After overexpression of PLUNC in HNE2 cells, decreased ATF3 and PD-L1 expression was confirmed by western blot and immunofluorescence. (E–H) After knocking down of PLUNC in 5-8F and HNE2 cells, upregulated ATF3 and PD-L1 expression was confirmed by western blot and immunofluorescence. (I, J) After the cells were treated with PLUNC-shRNA and/or XAV-939, the expressions of β-catenin, phospho–glycogen synthase kinase 3β (p–GSK-3β), ATF3, and PD-L1 were verified by western blot. NC, negative control; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
DDX17 interacts with β-catenin in NPC cells
However, the mechanism by which PLUNC downregulated β-catenin remains unclear. In 5-8F and HNE2 cells, PLUNC cannot bind directly to β-catenin, so we speculated that PLUNC may regulate β-catenin through another mechanism. The RNA helicase DDX17 is a member of a large family of highly conserved proteins that are involved in gene-expression regulation [27]. DDX17 acts as a co-activator or co-inhibitor of transcription factors of cell differentiation [28]. We explored the association of DDX17 with β-catenin in NPC cells. Just as seen in Fig. 5A–D, we confirmed DDX17 interactions with β-catenin by co-immunoprecipitation (Co-IP). Then, we demonstrated their co-localization by immunofluorescence experiments (Fig. 5E). Therefore, we further speculate whether PLUNC is related to the combination of DDX17 and β-catenin.
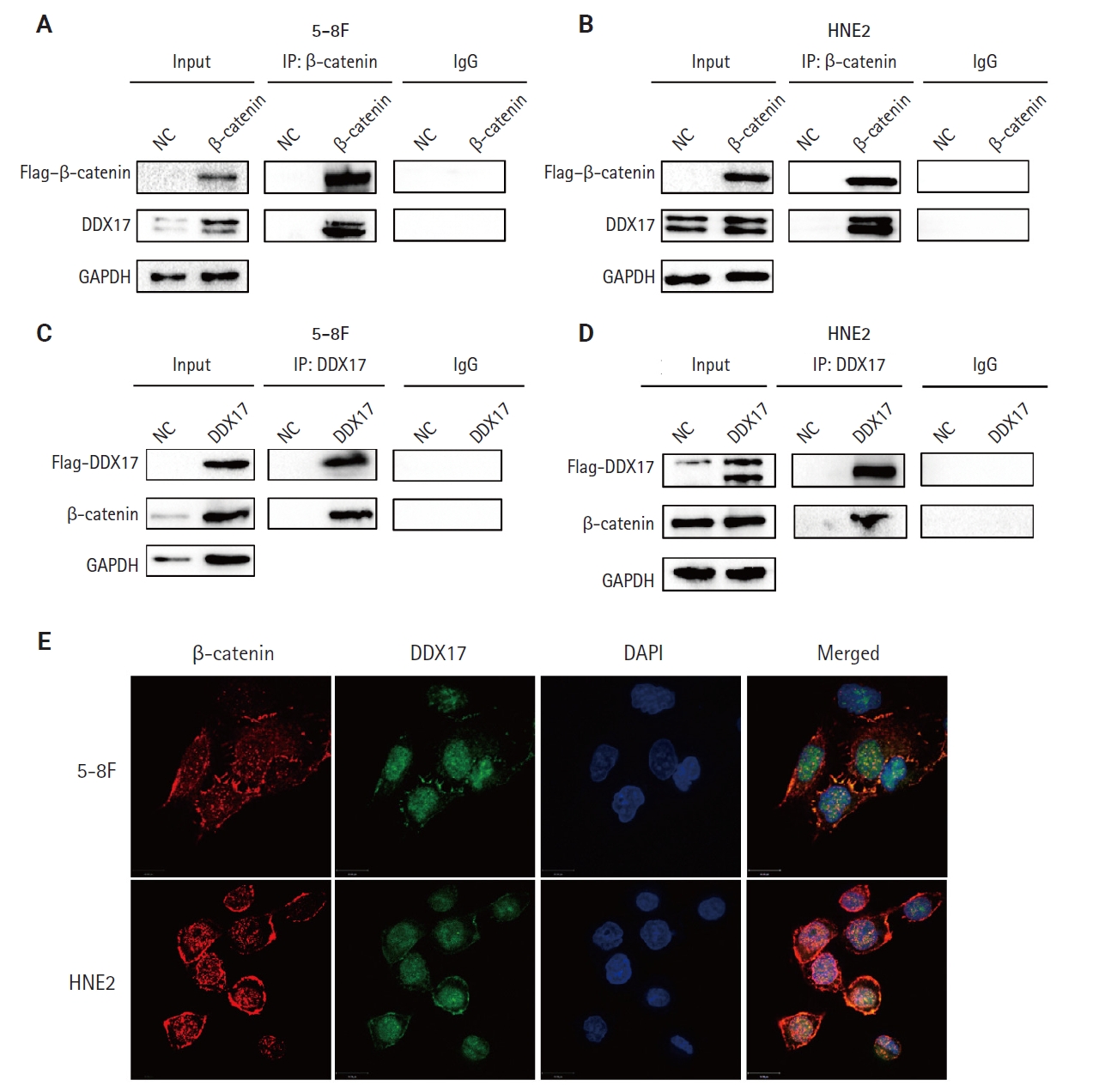
DEAD-box helicase 17 (DDX17) interacts with β-catenin in nasopharyngeal carcinoma cells. (A–D) Co-immunoprecipitation (Co-IP) of DDX17 and β-catenin in 5-8F and HNE2 cells. (E) Co-localization of DDX17 and β-catenin by immunofluorescence (β-catenin: red, DDX17: green). NC, negative control; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
PLUNC inhibits the interaction of DDX17 and β-catenin
In NPC cells, we confirmed the interactions between PLUNC and DDX17 by Co-IP (Fig. 6A, B). In addition, the co-localization of PLUNC and DDX17 was confirmed by immunofluorescence (Fig.6C, D). Therefore, we speculate that PLUNC and β-catenin compete to combine with DDX17. We co-transfected PLUNC, β-catenin, and DDX17 plasmids in 293T cells and successfully validated their expression (Fig. 6E). Then, we used the DDX17 antibody to co-precipitate PLUNC and β-catenin. We found that, following transfection with PLUNC, the interaction of DDX17 and β-catenin decreased, while, after transfection with β-catenin, the interaction of PLUNC and DDX17 decreased (Fig. 6F). In addition, through immunofluorescence experiments, we found that, after transfection with PLUNC, the co-localization of β-catenin and DDX17 decreased (Fig. 6G, H). Therefore, the results confirmed that PLUNC inhibits the interaction of DDX17 and β-catenin.
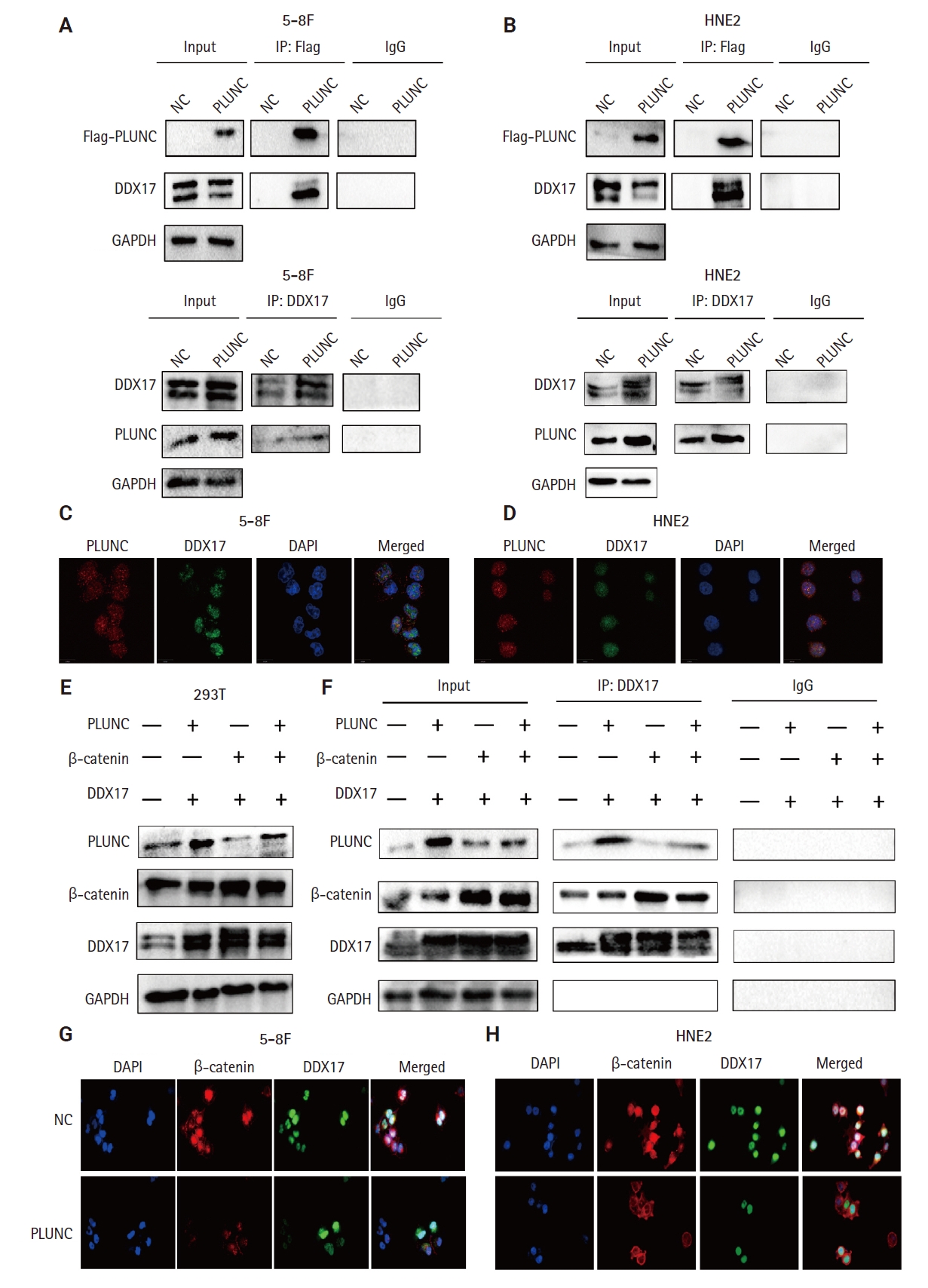
Palatal, lung, and nasal epithelial clone (PLUNC) inhibits the interaction of DEAD-box helicase 17 (DDX17) and β-catenin. (A, B) Co-immunoprecipitation (Co-IP) of DDX17 and PLUNC in 5-8F and HNE2 cells. (C, D) Co-localization of DDX17 and PLUNC by immunofluorescence (PLUNC: red, DDX17: green). (E) Successful co-transfection of PLUNC, β-catenin, and DDX17 plasmids in 293T cells was verified through western blot. (F) Co-IP confirmed that PLUNC inhibited the interaction of DDX17 and β-catenin. (G, H) After overexpression of PLUNC in 5-8F and HNE2 cells, the co-localization of DDX17 and β-catenin was reduced (β-catenin: red, DDX17: green). NC, negative control; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
PLUNC negatively correlates with DDX17 and β-catenin expression in NPC
In NPC tissue sections, we confirmed a negative correlation between PLUNC and the expressions of DDX17 and β-catenin. In NPC tissue, the expression of PLUNC was low, while those of DDX17 and β-catenin were high. However, in paracancer tissues, their expressions are exactly opposite (Fig. 7A). In addition, we present representative images with immunohistochemical scores in Supplementary Fig. S4.
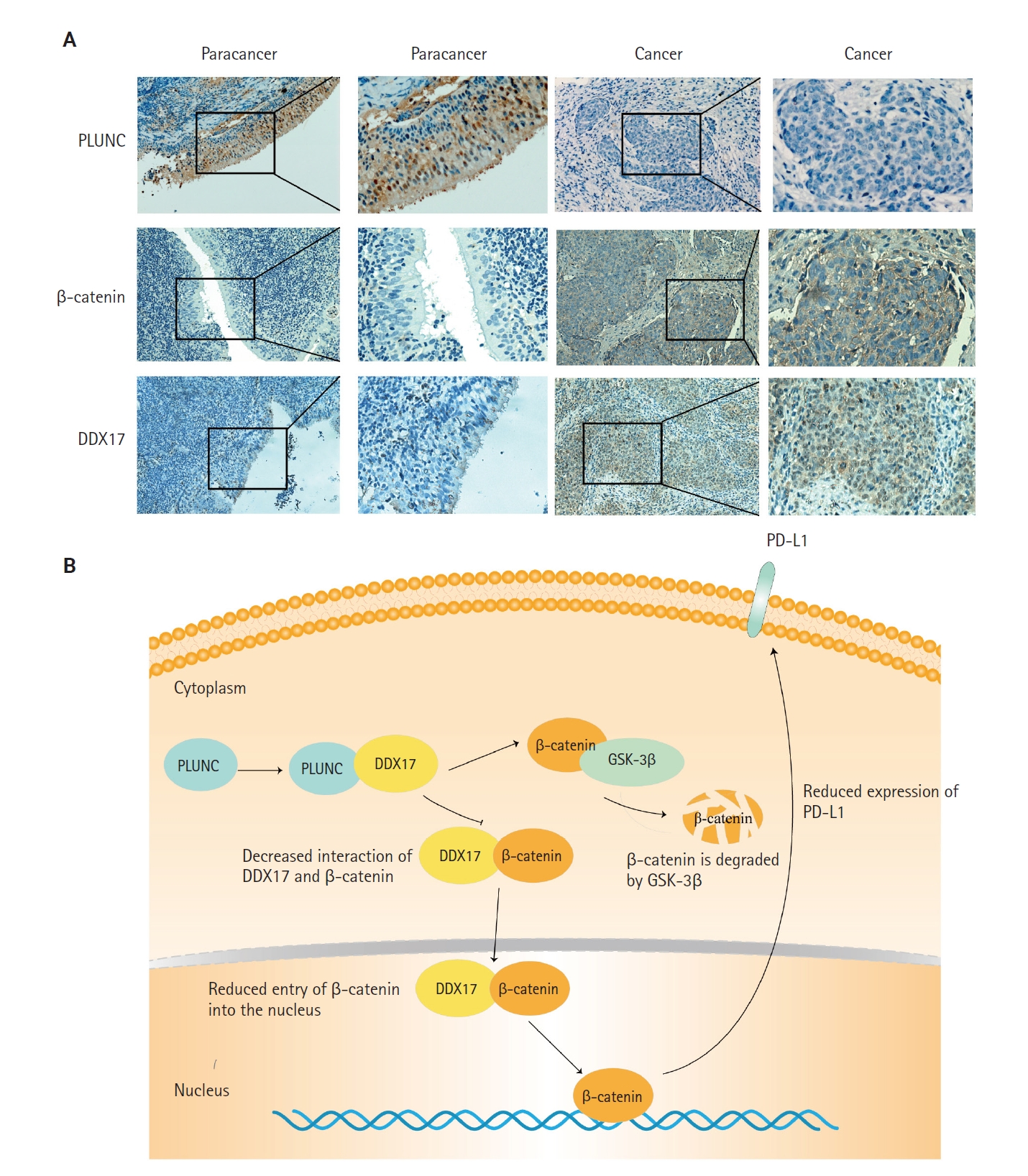
Palatal, lung, and nasal epithelial clone (PLUNC) negatively correlates with DEAD-box helicase 17 (DDX17) and β-catenin expression in nasopharyngeal carcinoma (NPC). (A) Immunohistochemical staining results of PLUNC, DDX17, and β-catenin in NPC and paracancer. (B) Research model diagram of PLUNC inhibiting programmed death-ligand 1 (PD-L1) expression. GSK-3β, glycogen synthase kinase 3β.
Finally, we concluded that the interaction of PLUNC and DDX17 inhibited the interaction of β-catenin and DDX17, thereby promoting the degradation of β-catenin by GSK-3β. However, the reduction of β-catenin's entry into the nucleus inhibited the expression of PD-L1, ultimately leading to a reduced expression of PLUNC (Fig. 7B).
DISCUSSION
EBV is closely related to the occurrence and development of NPC. It had been reported that EBV could upregulate the expression of PD-L1 in NPC cells [29-31]. Therefore, immunotherapy targeting PD-1/PD-L1 has become a new strategy for the treatment of NPC [32,33]. Our results indicated that high expression of PD-L1 is a risk factor for poor prognosis in patients with NPC. Therefore, inhibiting the expression of PD-L1 will benefit patients with NPC.
PLUNC was originally identified in the trachea and bronchi [34]. It was originally called PLUNC, and its gene product was later renamed short PLUNC 1 (SPLUNC1), also known as SPURT, LUNX, NASG, or BPIFA1. The secreted protein encoded by SPLUNC1 is abundant in human tracheobronchial secretions and saliva [35], nasal lavage [36], apical secretions of cultured human tracheobronchial epithelial cells [37,38], and specific granules of human neutrophils [39]. Previous research results from our group show that the poor prognosis of NPC patients closely relates to the low expression of PLUNC, which may represent a candidate molecular marker for early diagnosis of NPC [14]. In recent years, immunotherapy has become a promising treatment for NPC and has made great progress [40]. Upregulation of the PD-1/PD-L1 pathway is one of the possible mechanisms of immune-evasion in EBV-associated NPC [41]. PD-1/PD-L1 immune checkpoint inhibitors were approved in China in 2021 for the treatment of relapsed or metastatic NPC. In our study, PLUNC could downregulate the expression of PD-L1 in NPC.
Under physiological conditions, Wnt/β-catenin signaling is turned off. In the absence of Wnt signaling, a protein complex formed by AXIN, adenomatous polyposis coli (APC), serine/threonine kinase GSK-3β, CK1, and E3 ubiquitin ligase β-trcp degrades β-catenin [42]. Abnormal activation of Wnt/β-catenin signaling is common in tumors. When Wnt signaling is activated, GSK-3β is deactivated after phosphorylation, which leads to disassembly of the Axin/GSK3β/APC complex, thereby inhibiting the phosphorylation and degradation of β-catenin, and dephosphorylated β-catenin could be transported to the nucleus to regulate target genes [43]. Studies have shown that nuclear β-catenin can bind to the CD274 promoter region, promoting PD-L1 transcription and upregulating PD-L1 expression in nasopharyngeal carcinoma cells [44]. We also confirmed the expression of β-catenin in the nucleus and cytoplasm (Supplementary Fig. S3E, F). The results indicated that PLUNC could inhibit the expression of β-catenin in the nucleus and cytoplasm. However, the entry of β-catenin into the nucleus is reduced, leading to a decrease in the transcription of PD-L1 and ultimately downregulating the expression of PD-L1.
DDX17 (DEAD Asp-Glu-Ala-Asp) box helicase 17, also known as p72, is a typical member of the DEAD box family and is of interest for its role as an RNA helicase [45]. Among the DEAD-box RNA-binding proteins, DDX17 is a well-characterized cofactor of the Drosha/DGCR8 microprocessor, which mediates the recognition and processing of primary miRNAs (pri-miRNAs) to become mature miRNAs [46-48]. In addition, more and more studies are showing that DDX17 acts as a transcription factor-related protein that is associated with the function of transcription factors such as NFAT5, histone deacetylase 1, and SOX [49-52]. The abnormal expression of DDX17 could promote progression of various tumors [28,53,54]. Furthermore, DDX17 exerts its powerful oncogenic effects by acting as a co-activator or co-repressor of transcription factors, such as estrogen receptor α [55]. DDX17 could interact with β-catenin, while PLUNC could inhibit the β-catenin pathway. Therefore, we doubted whether PLUNC could regulate β-catenin through DDX17. Our experimental results also showed that PLUNC can directly interact with DDX17 and inhibited the interaction of DDX17 with β-catenin. After PLUNC knockdown, the expressions of both PD-L1 and β-catenin were increased.
ATF3 is a transcription factor that plays vital roles in modulating metabolism, immunity, and oncogenes that has sparked intense attention [56]. ATF3 could participate in the immune-regulation of the body and affect the occurrence, development, and treatment of tumors. Our results showed that β-catenin could upregulate ATF3 and PD-L1, while PLUNC can inhibit the Wnt/β-catenin pathway, thereby inhibiting the expressions of ATF3 and PD-L1. However, our experimental results showed that ATF3 was indeed regulated by β-catenin, while PD-L1 was indeed regulated by ATF3, but whether β-catenin directly regulates PD-L1 expression or relies on ATF3 to regulate PD-L1 still requires further experimental verification.
In conclusion, this study confirmed that PLUNC could downregulate the expression of PD-L1 in NPC by inhibiting the interaction of DDX17/β-catenin. This will provide new ideas for the immunotherapy of NPC.
Supplementary Information
The Data Supplement is available with this article at https://doi.org/10.4132/jptm.2024.11.27.
Notes
Ethics Statement
All procedures performed in the current study were approved by Xiangya Hospital of Central South University ethics committee (reference number: 202207389 Date:2022.07.05) in accordance with the 1964 Helsinki declaration and its later amendments. All specimens in this study had obtained informed consent from the patients.
Availability of Data and Material
The datasets generated or analyzed during the study are available from the corresponding author on reasonable request.
Code Availability
Not applicable.
Author Contributions
Conceptualization: RF, YG, MC, WZ. Formal analysis: JZ. Funding acquisition: WZ. Investigation: YG, MC, YL, QL, ZT, SJ, LS, SF. Supervision: XL, WX, GL, WZ. Writing—original draft: RF. Writing—review & editing: RF, WZ. Approval of final manuscript: all authors.
Conflicts of Interest
The authors declare that they have no potential conflicts of interest.
Funding Statement
This work was funded by the National Natural Science Foundation of China (Grant No. 81672688) and the Hunan Province Natural Science Foundation (Grant No. 2020JJ4780).
Acknowledgments
We thank The Third Xiangya Hospital of Central South University and The Cancer Research Institute of Central South University for its technical support.