Long Non-coding RNA HOTAIR Expression in Diffuse Large B-Cell Lymphoma: In Relation to Polycomb Repressive Complex Pathway Proteins and H3K27 Trimethylation
Article information
Abstract
Background
A long non-coding RNA hox transcript antisense intergenic RNA (HOTAIR) is involved in epigenetic regulation through chromatin remodeling by recruiting polycomb repressive complex 2 (PRC2) proteins (EZH2, SUZ12, and EED) that induce histone H3 trimethylation at lysine 27 (H3K27me3). Deregulation of c-MYC and interaction between c-MYC and EZH2 are well known in lymphomagenesis; however, little is known about the expression status of HOTAIR in diffuse large B-cell lymphomas (DLBCLs).
Methods
The expression status of PRC2 (EZH2, SUZ12, and EED), H3K27me3, c-MYC, and BCL2 was analyzed using immunohistochemistry (n = 231), and HOTAIR was investigated by a quantification real-time polymerase chain reaction method (n = 164) in DLBCLs.
Results
The present study confirmed the positive correlation among PRC2 proteins, H3K27me3, and c-MYC in DLBCLs. Expression level of HOTAIR was also positively correlated to EZH2 (p < .05, respectively). Between c-MYC and HOTAIR, and between c- MYC/BCL2 co-expression and HOTAIR, however, negative correlation was observed in DLBCLs (p < .05, respectively). High level of H3K27me3 was determined as an independent prognostic marker in poor overall survival (hazard ratio, 2.0; p = .023) of DLBCL patients. High expression of HOTAIR, however, was associated with favorable overall survival (p = .004) in the univariate analysis, but the impact was not significant in the multivariate analysis. The favorable outcome of DLBCL with HOTAIR high expression levels may be related to the negative correlation with c- MYC expression or c-MYC/BCL2 co-expression.
Conclusions
HOTAIR expression could be one of possible mechanisms for inducing H3K27me3 via EZH2-related PRC2 activation, and induced H3K27me3 may be strongly related to aggressive DLBCLs which show poor patient outcome.
Long non-coding RNAs (lncRNAs) are molecules longer than 200 nucleotides that are not translated into proteins. The role of lncRNAs is mostly unknown. Recently, several types of cancer-related lncRNAs have been identified and studied in the field of translational research [1-4]. According to recent studies, some lncRNAs are involved in the epigenetic regulation of protein coding genes. Hox transcript antisense intergenic RNA (HOTAIR) is one of most actively studied lncRNAs. Overexpression of HOTAIR is known to occur in various solid tumors of esophagus, stomach, colon, liver, pancreas, lung, and breast and is related to poor prognosis in those tumors [3,5]. HOTAIR is located within the homeobox C (HOXC) gene cluster on chromosome12 and is co-expressed with the HOXC genes. HOTAIR is involved in chromatin remodeling through recruiting polycomb repressive complex 2 (PRC2; enhancer of zeste homolog 2 [EZH2], suppressor of zeste 12 homolog [SUZ12], and embryonic ectoderm development [EED]) and then inducing histone modification such as histone H3 trimethylation at lysine 27 (H3K27me3) at the promoter site of protein-coding genes [1-4]. Chromatin remodeling and gene regulation via histone modification of the polycomb repressive complex is known to function in the development of embryonic stem cells as well as the development of many types of cancers including hematologic malignancies [6,7].
In hematologic malignancies, especially in diffuse large B cell lymphomas (DLBCLs) and follicular lymphomas that are the most predominant lymphoma subtype, the deregulation of EZH2 methyltransferase is well known. In our previous study, the high level of global H3K27me3 in DLBCL was associated with poor patient prognosis [8]. From these findings, it was suggested that HOTAIR might be involved in the PRC2-associated induction of H3K27me3 in DLBCLs; however, an association with HOTAIR and DLBCL has not yet been described. In the present study, the expression status of HOTAIR was investigated in DLBCL, and the association with PRC2 and H3K27me3 was analyzed.
MATERIALS AND METHODS
Patients and clinical data
A total of 231 cases of DLBCL treated with R-CHOP (rituximab, cyclophosphamide, doxorubicin, vincristine, and prednisone) or R-CHOP–like (with or without radiotherapy or surgery) chemotherapy were selected for the study. Cases were retrieved from the archival files from the Department of Pathology, Severance Hospital, from 2005 to 2011. All cases were independently reviewed by two pathologists (S.O.Y. and S.H.K.) based on current World Health Organization criteria [9], and discordant cases were consulted to other expert hematopathologists. In HOTAIR expression analysis, 164 cases were selected from the above 231 cases and investigated after quality assessment of extracted RNA. Clinical data were obtained from medical records. All study protocols were performed according to the ethical guidelines of the ‘‘World Medical Association Declaration of Helsinki–Ethical Principles for Medical Research Involving Human Subjects.’’ This study was approved by the Institutional Review Board of Severance Hospital.
Analysis for HOTAIR expression
Formalin fixed paraffin embedded (FFPE) tissue sections were prepared and stained with hematoxylin and eosin, and then the tumor areas were confirmed and marked under the microscope. The marked areas mainly contained packed tumor cells, and the stromal component was less than 10% of the marked area. The unstained slides of FFPE tissues were prepared after dissecting FFPE tissue blocks at 10-mm thickness using a microtome, and the marked area was scraped using a scalpel blade. Generally, three slices of tissue section per case were used for RNA extraction. Total RNA was isolated using an RNeasy FFPE Kit (Qiagen, Hilden, Germany) according to the supplier’s instructions. Extracts of RNA were verified by measuring the ratios ofA260/A280 and A260/A230 with a ND-1000 NanoDrop spectrophotometer (NanoDrop, Wilmington, DE, USA). Reverse transcription was performed using a QuantiTect Reverse Transcription kit (Qiagen). The expression patterns of HOTAIR were assayed by relative quantification using expression of the house-keeping gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Primers for HOTAIR and GAPDH were as follows: HOTAIR (forward, 5'-AGCCAGAGGAGGGAAGAGAG-3'; reverse, 5'-TCCCGTTCCCTAGATTTTCC-3') and GAPDH (forward, 5'-CAAATTCCATGGCACCGTCA-3'; reverse, 5'-ATCGCCCCACTTGATTTTGG-3'). Primers of HOTAIR were designed to detect all three transcript variants (transcript variant 1, 3, and 2). In brief, a 20 μL mixture containing 1.0 μL of cDNA, power SYBR Green PCR Master Mix (Applied Bio-systems, Carlsbad, CA, USA), 1.0 μL of 10 pmol/μL forward primer, 1.0 μL of 10 pmol/μL reverse primer, and 7.0 μL of tertiary distilled water was prepared. Amplification was performed using a Step One Plus Real-Time PCR instrument (Applied Biosystems) under the following conditions: denaturation at 94°C for 10 minutes, followed by 35 cycles of 94°C for 15 seconds, 55°C for 30 seconds, and 72°C for 30 seconds (fluorescence signal acquisition was performed at this phase). Immediately after amplification, melting curve analysis was performed for amplicon verification. All samples were analyzed in triplicate to confirm reproducibility. Using the Step One Plus Real-Time PCR System software ver. 2.1 (Applied Biosystems), the threshold cycle (Ct, beginning of the polymerase chain reaction exponential phase) value of amplified HOTAIR was normalized versus that of amplified GAPDH (2-dCt). After quality assessment of extracted RNA and expression analysis of the housekeeping gene GAPDH, 164 cases were finally selected for the analysis of HOTAIR expression. Normal palatine tonsil tissue was obtained from 10 cancer-free individuals and used as age-matched cancer-free controls. The cutoff value for high expression of HOTAIR (HOTAIRhigh) was determined at the uppermost value among those of normal control tonsil tissues.
Tissue microarray preparation, immunohistochemistry, and analysis
The hematoxylin and eosin slides were reviewed and two representative core tissues of the tumor area were selected in each case. Core tissues (3 mm in diameter) were taken from the individual donor blocks and arranged in new recipient tissue microarray paraffin blocks using a trephine apparatus. Immunohistochemical staining and in situ hybridization were performed on 4-μm tissue microarray tissue sections. Immunohistochemistry of EZH2 (1:100, Invitrogen, Carlsbad, CA, USA), SUZ12 (1:50, Abcam, Cambridge, UK), EED (1:1,000, Abcam), H3K27me3 (1:100, clone C36B11, Cell Signaling Technology, Beverly, MA, USA) was performed using the Ventana BenchMark XT Autostainer (Ventana Medical Systems, Tucson, AZ, USA). Immunohistochemistry for c-MYC (1:50, clone Y69, Abcam), BCL2 (1:50, Novocastra, Newcastle, UK), CD10 (1:100, Novocastra), BCL6 (RTU, Novocastra), and MUM1 (1:200, Cell Marque, Rocklin, CA, USA) was performed using the LEICA BOND-III Autostainer (Leica Biosystems, Newcastle Upon Tyne, UK). For in situ hybridization for Epstein-Barr virus (EBV), the INFORM EBER probe (Ventana Medical Systems) was used with the Ventana BenchMark XT Autostainer (Ventana Medical Systems) and ISH iVIEW Blue Detection kit (Ventana Medical Systems).
For EZH2, SUZ12, EED, and H3K27me3, staining intensity (0, no staining to weak; 1, moderate to strong) and proportion of positive tumor cell nuclei (0, <10%; 1, ≥10% and <75%; 2, ≥75%) were semiquantitatively graded as in the previous study [8]. Based on the intensity multiplied by proportion, the protein expression was scored as low (0), intermediate (1), or high level (2). For CD10, BCL6, and MUM1, the positive cutoff value was determined according to the Hans classification criteria [10] and was considered positive if ≥30% of the tumor cells showed nuclear immunoreactivity for BCL6 and MUM1, and if ≥30% of cells showed membranous reactivity for CD10. Determination of the germinal center B-like (GCB) or non-GCB phenotype was based on the Hans algorithm 10.
For c-MYC and BCL2, staining intensity (0, no staining to weak; 1, moderate to strong) and areas of positive tumor cell nuclei by 10% increments (1 of <10% to 10 of 90%–100%) were semiquantitatively graded. The cutoff value for high expression was determined by log-rank tests (Mantel-Cox) for overall survival as in the previous report, and the threshold value was determined for c-MYC at a score ≥4 (≥40% of tumor cells with moderate to strong expression), and BCL2 at a score ≥7 (≥70% of tumor cells with moderate to strong expression) [11,12]. For EBV in situ hybridization, the threshold value was determined when ≥ 10% of the tumor cells showed moderate to strong nuclear expression.
The features of expression of polycomb repressive complex proteins (EZH2, SUZ12, and EED), H3K27me3, c-MYC, and BLC2 in HOTAIRhigh or HOTAIRlow cases are presented in Fig. 1.
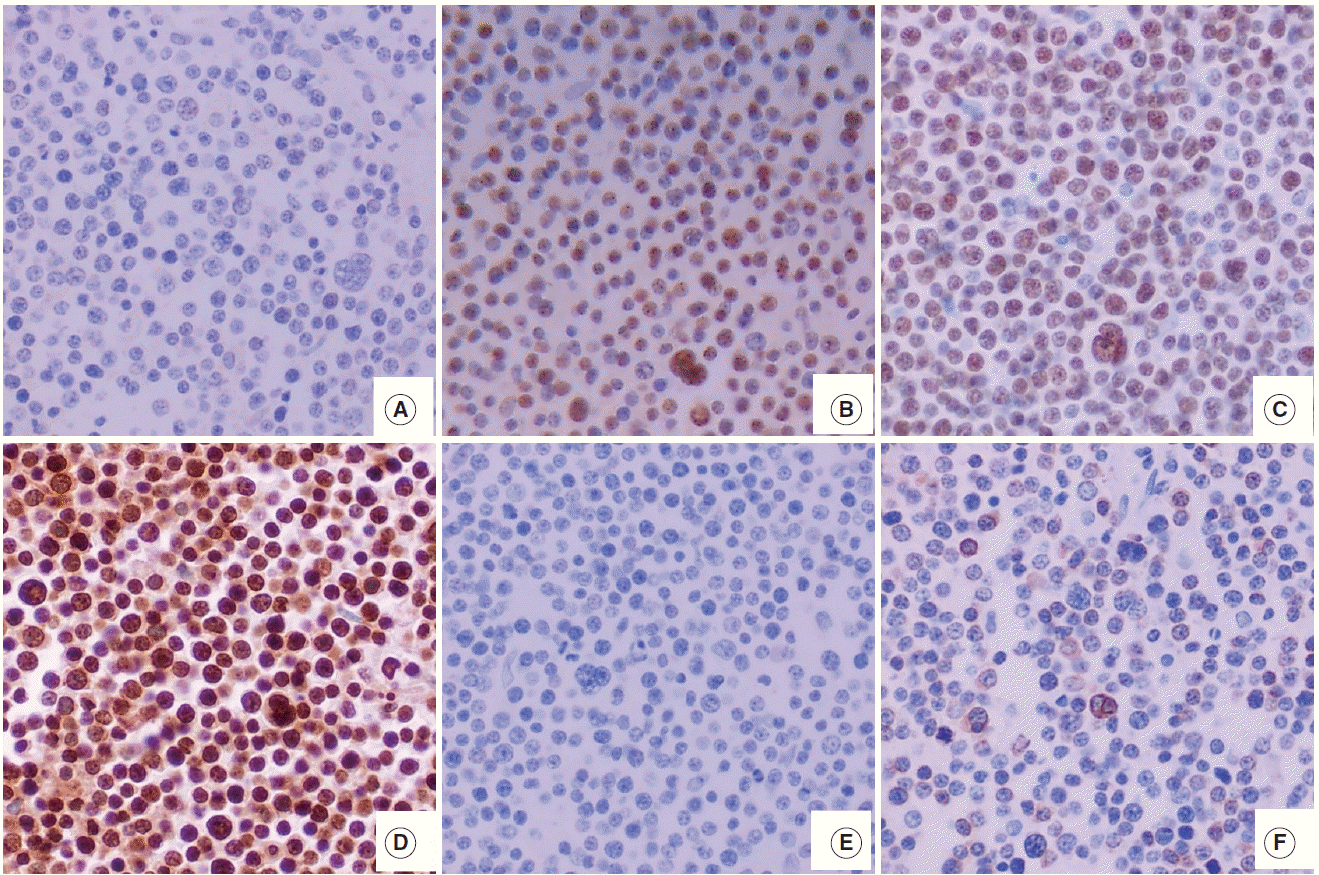
A case of DLBCL with HOTAIRhigh (A–F) shows the various expression pattern of c-MYC (A), EZH2 (B), SUZ12 (C), EED (D), global H3K27me3 (E), and BCL2 (F). In a DLBCL case of HOTAIRhigh (A–F), many PRC2 protein (EZH2, SUZ12, and EED)–positive tumor cells show no expression of c-MYC and no mark of H3K27me3. BCL2 was negative in this case. DLBCL, diffuse large B-cell lymphoma; HOTAIR, hox transcript antisense intergenic RNA; EZH2, enhancer of zeste homolog 2; SUZ12, suppressor of zeste 12 homolog; EED, embryonic ectoderm development; PRC2, polycomb repressive complex 2.
Statistical analysis
The t test and chi-square test were used to analyze the differences between the variables examined. Overall survival times were measured from the date of lymphoma diagnosis to the date of death or last follow-up visit. Patient survival rates were determined using the Kaplan-Meier method, and the differences in survival rates were compared using the log-rank test. Multivariate analysis was performed using the Cox proportional hazards model. A two-sided p-value<.05 was considered to be statistically significant. When a two-sided p-value was ≤.05 and <.10, a trend toward statistical significance was considered. All statistical analyses were carried out using SPSS software ver. 20.0 for Windows (IBM Corp., Armonk, NY, USA).
RESULTS
The relations among PRC2 proteins, H3K27me3, c-MYC, BCL2, and HOTAIR
The DLBCL cases frequently showed high expression of PRC2 proteins and H3K27me3 (28.9%, 47.1%, 74.3%, and 35.7% for EZH2high, SUZ12high, EEDhigh, and H3K27me3high, respectively). High expression of all PRC2 proteins (EZH2high/SUZ12high/EEDhigh) was noted in 16.1% of cases (Fig. 2A). The expression of H3K27me3 was positively correlated to PRC2 markers. When compared to cases of H3K27me3low, those of high H3K27me3 expression more frequently showed high expression of EZH2 (41.2% vs 22.2%, p=.003), SUZ12 (62.5% vs 43.0%, p=.053), EED (88.6% vs 69.7%, p=.026), and all PRC2 (EZH2high/SUZ12high/EEDhigh) (32.3% vs 11.3%, p=.005). These results are summarized in Fig. 2B.
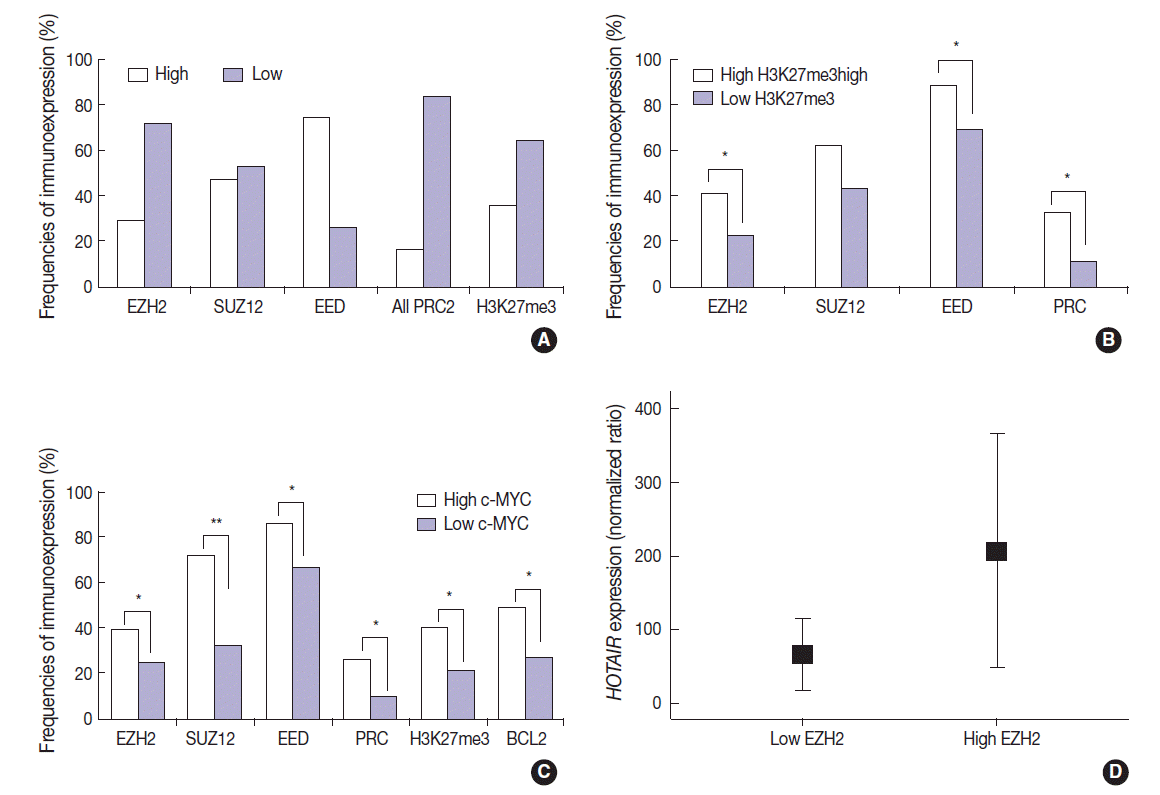
(A) The DLBCL cases frequently show high expression of PRC2 proteins (EZH2, SUZ12, and EED), high expression of all PRC2 proteins (EZH2high/SUZ12high/EEDhigh), and high H3K27me3 level. (B) When compared to cases without high H3K27me3 expression (low H3K27me3), those with high H3K27me3 expression more frequently show high expression of EZH2, SUZ12, EED, and all PRC2 (EZH2high/SUZ12high/EEDhigh). (C) When compared to cases without high c-MYC expression (low c-MYC), those with high c-MYC expression more frequently show high expression of EZH2, SUZ12, EED, all PRC2 (EZH2high/SUZ12high/EEDhigh), H3K27me3, and BCL2. (D) The expression level of HOTAIR is significantly higher in cases with high EZH2 expression than those without high EZH2 expression (low EZH2). DLBCL, diffuse large B-cell lymphoma; HOTAIR, hox transcript antisense intergenic RNA; EZH2, enhancer of zeste homolog 2; SUZ12, suppressor of zeste 12 homolog; EED, embryonic ectoderm development; PRC2, polycomb repressive complex 2. *p < .05, **p < .001.
The expression of c-MYC was positively correlated to PRC2 markers and H3K27me3. When compared to cases of c-MYClow, those of high c-MYC expression more frequently showed high expression of EZH2 (39.1% vs 24.6%, p=.036), SUZ12 (72.2% vs 32.1%, p<.001), EED (85.7% vs 66.7%, p=.011), all PRC2 (EZH2high/SUZ12high/EEDhigh) (26.4% vs 9.5%, p=.009), and H3K27me3 (40.6% vs 21.2%; p=.004). BCL2 was positively correlated with c-MYC (49.3% vs 26.8%; p=.002). These results are summarized in Fig. 2C. BCL2 expression showed no significant correlation with the tested PRC2-related markers.
The expression level of HOTAIR was significantly higher in cases of EZH2high than those of EZH2low (mean ratio value, 207 vs 66; p=.027) (Fig. 2D). For other markers, no statistical significance was observed according to HOTAIR expression level.
HOTAIR expression in in relation to clinicopathological variables of DLBCL
In the present study, 23.8% of cases (39/164) showed high HOTAIR expression levels (HOTAIRhigh). In the correlation analysis between HOTAIRhigh and clinicopathological characteristics of DLBCL (Table 1), high expression of c-MYC protein was less frequent in HOTAIRhigh than in HOTAIRlow (p=.037). The rate of co-expression of c-MYC and BCL2 was significantly lower in HOTAIRhigh than in HOTAIRlow (p=.015). Other clinicopathological factors including EBV (p=.660) and Hans classification (p=.746) showed no significant correlation with HOTAIR expression (Table 1).
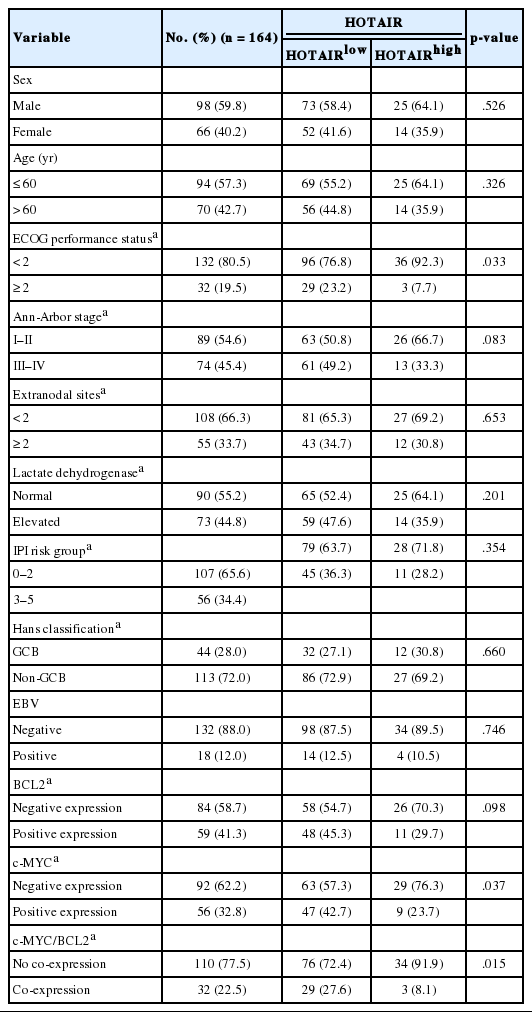
Clinicopathological characteristics, relation to outcome, and association with HOTAIR level in DLBCL patients
Clinical significance of RNA and protein expression
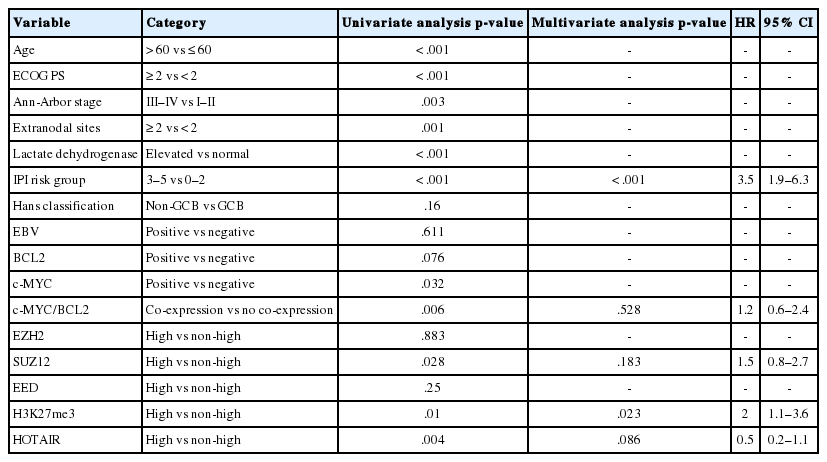
In univariate analysis for overall survival of DLBCL patients (Table 2), the known prognostic factors including age >60 (p<.001), Eastern Cooperative Oncology Group performance status ≥2 (p<.001), elevated lactate dehydrogenase (p<.001), extranodal sites ≥2 (p=.001), Ann-Arbor stage III–IV (p=.003), and International Prognostic Index (IPI) risk ≥3 (p<.001) showed significant association with reduced overall survival rate. Expression of c-MYC and co-expression of c-MYC/BCL2 were also related to inferior overall survival rate (p=.032 and p=.006, respectively). Among the PRC pathway markers (EZH2, SUZ12, EED, and H3K27me3), SUZ12high and H3K27me3high were related to inferior overall survival rate (p=.028 and p=.010, respectively) (Fig. 3A, B). HOTAIRhigh, however, was significantly related to superior overall survival rate than HOTAIRlow (p=.004) (Fig. 3C).
Cox proportional hazard analyses of overall survival
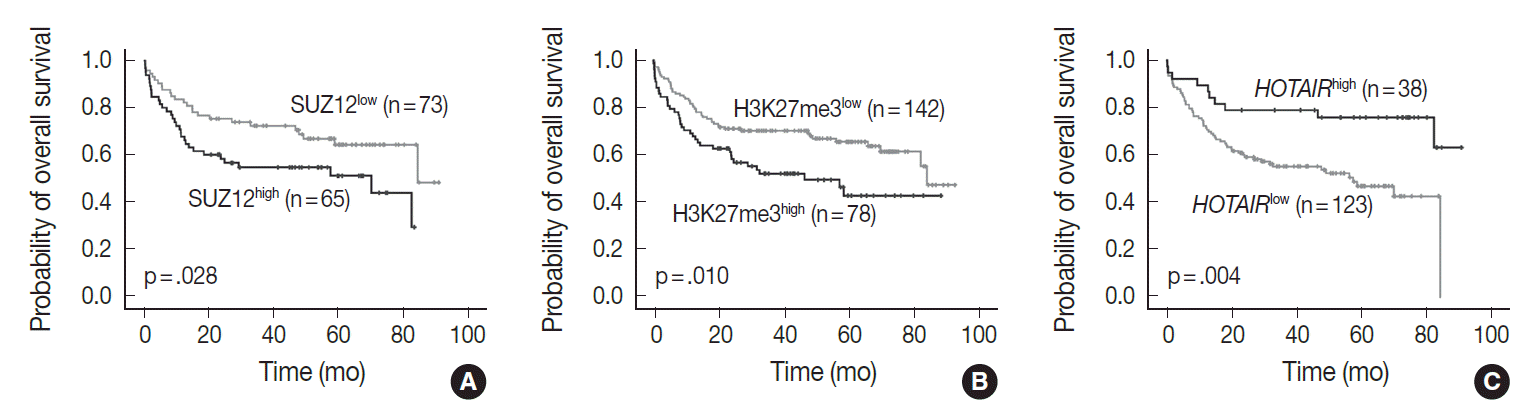
Overall survival based on SUZ12 (A), H3K27me3 (B), and HOTAIR expression (C). High expression of SUZ12 (SUZ12high) and H3K27me3 (H3K27me3high) is related to inferior overall survival, while high expression of HOTAIR (HOTAIRhigh) is correlated with longer overall survival than low levels of HOTAIR expression (HOTAIRlow). SUZ12, suppressor of zeste 12 homolog; HOTAIR, hox transcript antisense intergenic RNA.
In multivariate analysis (Table 2), H3K27me3high revealed an independent effect on poor overall survival (hazard ratio, 2.0; p=.023). IPI risk ≥3 was still determined as an independent prognostic factor (hazard ratio, 3.5; p<.001), while HOTAIRhigh showed a tendency to be related to improved survival with marginal significance (hazard ratio, 0.5; p=.086).
DISCUSSION
In our previous study, high level of global H3K27me3 was found to be a negative prognostic indicator in patients with DLBCL [8]. This subsequent study aimed to explore the role of HOTAIR, possibly functioning via induction of H3K27me3. We found the positive correlation between PRC2 proteins, global H3K27me3 levels, and c-MYC in DLBCL as expected.
In the present study, the positive correlation among PRC2 proteins (EZH2, SUZ12, and EED), global H3K27me3 levels, and c-MYC was also confirmed in DLBCL as expected. In addition, HOTAIR expression was related to EZH2 expression. These findings could support that the lncRNA HOTAIR may be involved in inducing H3K27me3 through recruiting polycomb repressive complex, the methyltransferase EZH2 and core accessory proteins, SUZ12 and EED.
Based on many other studies, interaction between c-MYC and PRC2 (EZH2, SUZ12, and EED) is well known in the tumorigenesis of various cancer types including lymphomas. Protein c-MYC interacts with EZH2 as well as SUZ12/EED, and they induce the histone modification of H3K27me3 on the promoter of target genes, which then represses gene expression. Many other studies have shown that EZH2 and c-MYC activate each other [13-15]. From the present findings, it could be also suggested that c-MYC may be the possible mechanism for inducing H3K27me3 via PRC2-related pathways in DLBCLs.
In other studies of solid tumors originating from various organs, high expression of HOTAIR showed a close association with poor prognosis [5]. In the present study, however, high levels of HOTAIR expression showed an association with good prognosis in DLBCL, although the impact was not significant in the multivariate analysis. There are no comparable reports to support the present findings because this is the first study for the expression status of HOTAIR in hematologic malignancies.
Although interaction between c-MYC and HOTAIR has not been established in hematologic malignancies, a close relationship between them may be plausible when considering the close association of PRC2 and HOTAIR, and PRC2 and c-MYC. However, expression of c-MYC showed a negative correlation with HOTAIR in the present study. One of the possible reason is autoregulation of c-MYC reported in one study; c-MYC represses itself via forming autoregulatory loops with EZH [2,13] therefore, HOTAIR might be involved in that process via EZH2. Whether HOTAIR expression is directly linked to the suppression of c-MYC or H3K27me3 should be investigated in further functional studies. In the HOTAIR-related suppression of c-MYC, PRC2-associated histone modification (H3K27me3) might be induced in the promoter region of c-MYC gene. For confirmation of this possible modulation, further study should be followed.
The favorable outcome of DLBCLs with high HOTAIR expression might be associated with the negative correlation with c-MYC and/or BCL2. Recent evidence has shown that co-expression of c-MYC and BCL2 proteins is associated with poor prognosis in DLBCL patients regardless of gene signature [11,12]. This was also observed in the present study in the univariate survival analysis. When the factor of HOTAIR expression was added in the multivariate analysis; however, the prognostic effect of coexpression of c-MYC and BCL2 became weak, and only the factor of high H3K27me3 level was important as well as the IPI risk score. H3K27me3, the chromatin modification status induced via changes of HOTAIR, c-MYC, or other various known and unknown mechanisms, seems to be the most important factor in determining the fate of disease aggressiveness of DLBCL. Though little is known about the key role of HOTAIR in the malignant lymphoma, HOTAIR might be involved in regulation of various genes in the lymphomagenesis. Further study should follow to determine the explicit mechanism among the sophisticated modulatory networks of c-MYC, PRC2, H3K27me3, and HOTAIR.
Recent evidence has shown that inhibition of EZH2 methyltransferase activity provides a potential target therapy for EZH2-deregulated lymphomas. In addition, the pharmacological inhibition of EZH2 activity results in a decrease of global H3K27me3 levels and a reactivation of silenced PRC2 gene targets, and it inhibits growth of DLBCL cells [16,17]. From these overall findings as well as the present findings, HOTAIR may also be used in a target therapy by modulating the c-MYC–EZH2/PRC2 loop in a subset of DLBCLs with a high level of H3K27me3.
In conclusion, we found frequent expression of PRC2 proteins and H3K27me3 and positive correlation between these proteins and c-MYC in DLBCLs. High expression of H3K27me3 was determined as an independent predictor of poor prognosis, however, HOTAIR was associated with favorable overall survival, which can be partly explained by negative correlation with c-MYC. LncRNA HOTAIR expression could be one of the possible mechanisms to be involved in aggressive behavior of DLBCL via induction of H3K27me3 and EZH2-related PRC2 activation.
Notes
Conflicts of Interest
No potential conflict of interest relevant to this article was reported.
Acknowledgements
This work was supported by grant no. 2014-01 from the Korean Medical Women’s Association.